

cAMP response element modulator α controls IL2 and IL17A expression during CD4 lineage commitment and subset distribution in lupus
- PMID: 23019580
- PMCID: PMC3478624
- DOI: 10.1073/pnas.1210129109
cAMP response element modulator α controls IL2 and IL17A expression during CD4 lineage commitment and subset distribution in lupus
Abstract
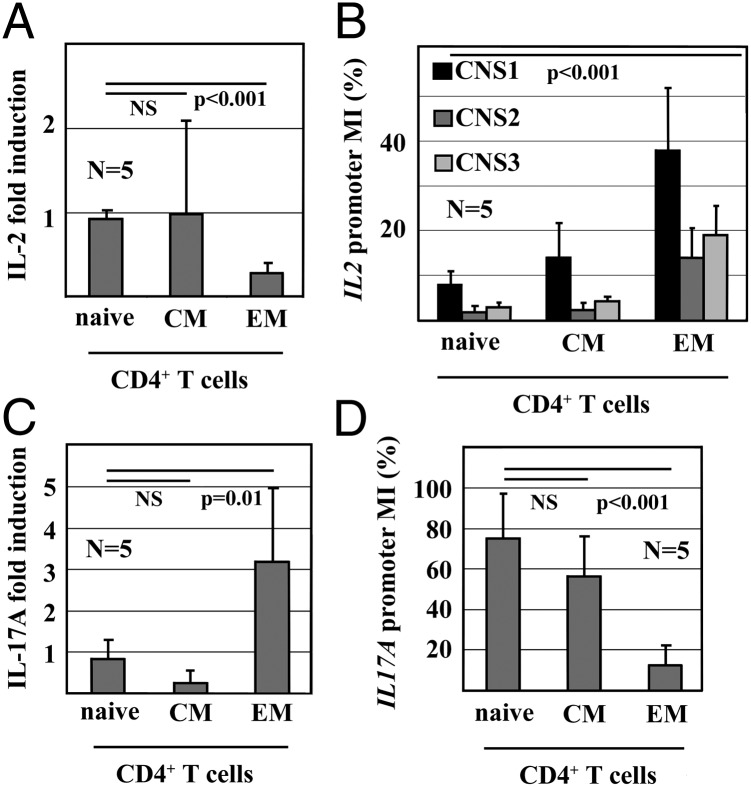
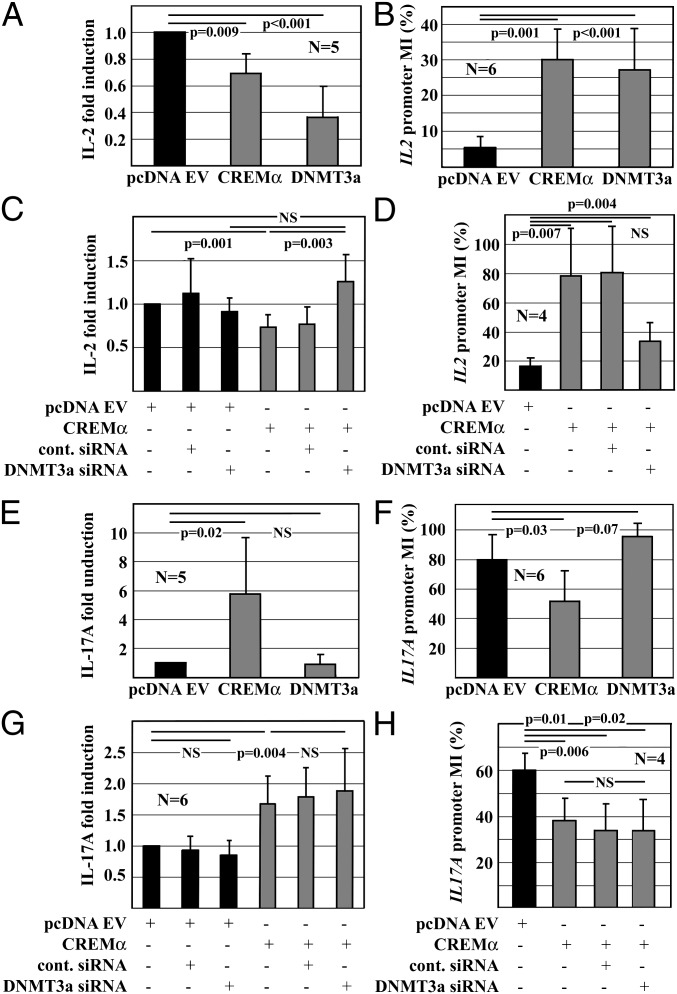
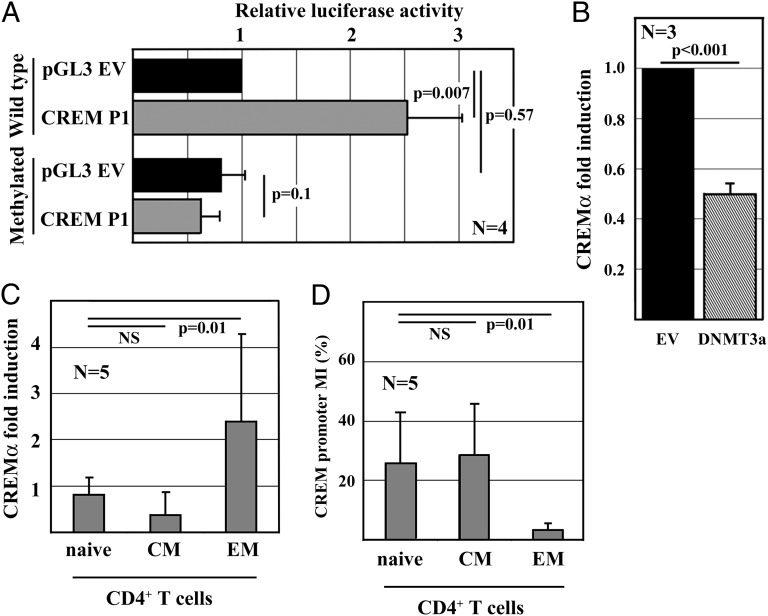
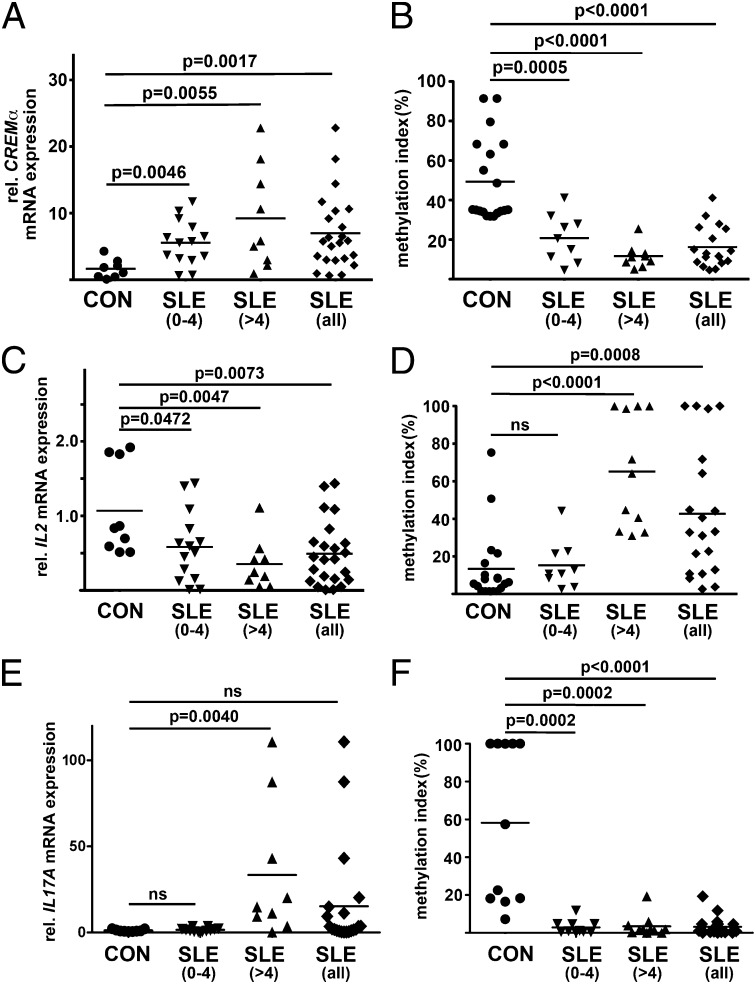
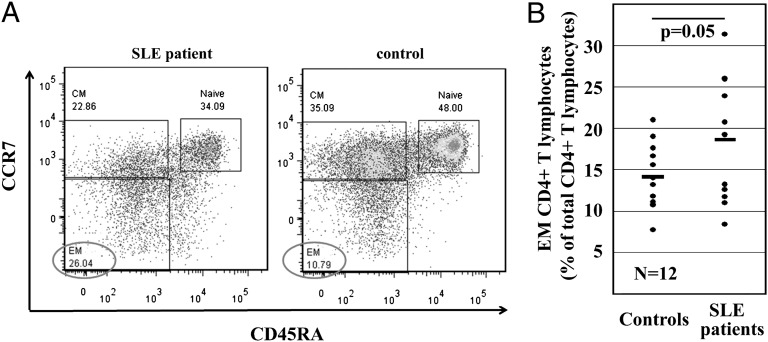
Appropriate expression of IL-2 plays a central role during the priming and differentiation of T cells. A tight balance between IL-2 and the effector cytokine IL-17A is essential for immune homeostasis. Epigenetic mechanisms have been documented as a key component of cytokine regulation during lineage commitment. The molecular mechanisms that induce chromatin remodeling are less well understood. We investigated epigenetic regulators that mediate the diametric expression of IL-2 and IL-17A in naive, central memory, and effector memory CD4(+) T cells. We demonstrate that cAMP response modulator (CREM)α contributes to epigenetic remodeling of IL2 in effector memory T cells through the recruitment of DNMT3a. CREMα also reduces CpG-DNA methylation of the IL17A promoter. CREMα expression is regulated at the epigenetic level by CpG-DNA methylation, which allows increased CREMα expression in effector memory CD4(+) T cells. T cells from patients with systemic lupus erythematosus (SLE) express increased levels of CREMα and exhibit a phenotype that is similar to effector memory CD4(+) T cells with epigenetically predetermined expression patterns of IL-2 and IL-17A. We conclude that CREMα mediates epigenetic remodeling of the IL2 and IL17A gene during T-cell differentiation in favor of effector memory T cells in health and disease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Increased Set1 binding at the promoter induces aberrant epigenetic alterations and up-regulates cyclic adenosine 5'-monophosphate response element modulator alpha in systemic lupus erythematosus.Clin Epigenetics. 2016 Nov 24;8:126. doi: 10.1186/s13148-016-0294-2. eCollection 2016. Clin Epigenetics. 2016. PMID: 27904655 Free PMC article.
-
cAMP-responsive element modulator (CREM)α protein induces interleukin 17A expression and mediates epigenetic alterations at the interleukin-17A gene locus in patients with systemic lupus erythematosus.J Biol Chem. 2011 Dec 16;286(50):43437-46. doi: 10.1074/jbc.M111.299313. Epub 2011 Oct 24. J Biol Chem. 2011. PMID: 22025620 Free PMC article.
-
Decreased SUV39H1 at the promoter region leads to increased CREMα and accelerates autoimmune response in CD4+ T cells from patients with systemic lupus erythematosus.Clin Epigenetics. 2022 Dec 20;14(1):181. doi: 10.1186/s13148-022-01411-7. Clin Epigenetics. 2022. PMID: 36536372 Free PMC article.
-
The cAMP responsive element modulator (CREM) is a regulator of CD4+ T cell function.Biol Chem. 2021 Aug 26;402(12):1591-1596. doi: 10.1515/hsz-2021-0249. Print 2021 Nov 25. Biol Chem. 2021. PMID: 34448385 Review.
-
[Interleukin-2 signaling pathway regulating molecules in systemic lupus erythematosus].Beijing Da Xue Xue Bao Yi Xue Ban. 2016 Dec 18;48(6):1100-1104. Beijing Da Xue Xue Bao Yi Xue Ban. 2016. PMID: 27987522 Review. Chinese.
Cited by
-
cAMP responsive element modulator: a critical regulator of cytokine production.Trends Mol Med. 2013 Apr;19(4):262-9. doi: 10.1016/j.molmed.2013.02.001. Epub 2013 Mar 13. Trends Mol Med. 2013. PMID: 23491535 Free PMC article. Review.
-
Calcium/Calmodulin Kinase IV Controls the Function of Both T Cells and Kidney Resident Cells.Front Immunol. 2018 Oct 1;9:2113. doi: 10.3389/fimmu.2018.02113. eCollection 2018. Front Immunol. 2018. PMID: 30333818 Free PMC article. Review.
-
T cells and autoimmune kidney disease.Nat Rev Nephrol. 2017 Jun;13(6):329-343. doi: 10.1038/nrneph.2017.34. Epub 2017 Mar 13. Nat Rev Nephrol. 2017. PMID: 28287110 Review.
-
Pathogenesis of Human Systemic Lupus Erythematosus: A Cellular Perspective.Trends Mol Med. 2017 Jul;23(7):615-635. doi: 10.1016/j.molmed.2017.05.006. Epub 2017 Jun 13. Trends Mol Med. 2017. PMID: 28623084 Free PMC article. Review.
-
Helios as a Potential Biomarker in Systemic Lupus Erythematosus and New Therapies Based on Immunosuppressive Cells.Int J Mol Sci. 2023 Dec 29;25(1):452. doi: 10.3390/ijms25010452. Int J Mol Sci. 2023. PMID: 38203623 Free PMC article. Review.
References
-
- Sallusto F, Lanzavecchia A. Human Th17 cells in infection and autoimmunity. Microbes Infect. 2009;11:620–624. - PubMed
-
- Zielinski CE, et al. Pathogen-induced human TH17 cells produce IFN-γ or IL-10 and are regulated by IL-1β. Nature. 2012;484:514–518. - PubMed
-
- Sallusto F, Lenig D, Förster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401:708–712. - PubMed
-
- Wilson CB, Rowell E, Sekimata M. Epigenetic control of T-helper-cell differentiation. Nat Rev Immunol. 2009;9:91–105. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
