

D-2-hydroxyglutarate produced by mutant IDH1 perturbs collagen maturation and basement membrane function
- PMID: 22925884
- PMCID: PMC3444730
- DOI: 10.1101/gad.198200.112
D-2-hydroxyglutarate produced by mutant IDH1 perturbs collagen maturation and basement membrane function
Abstract
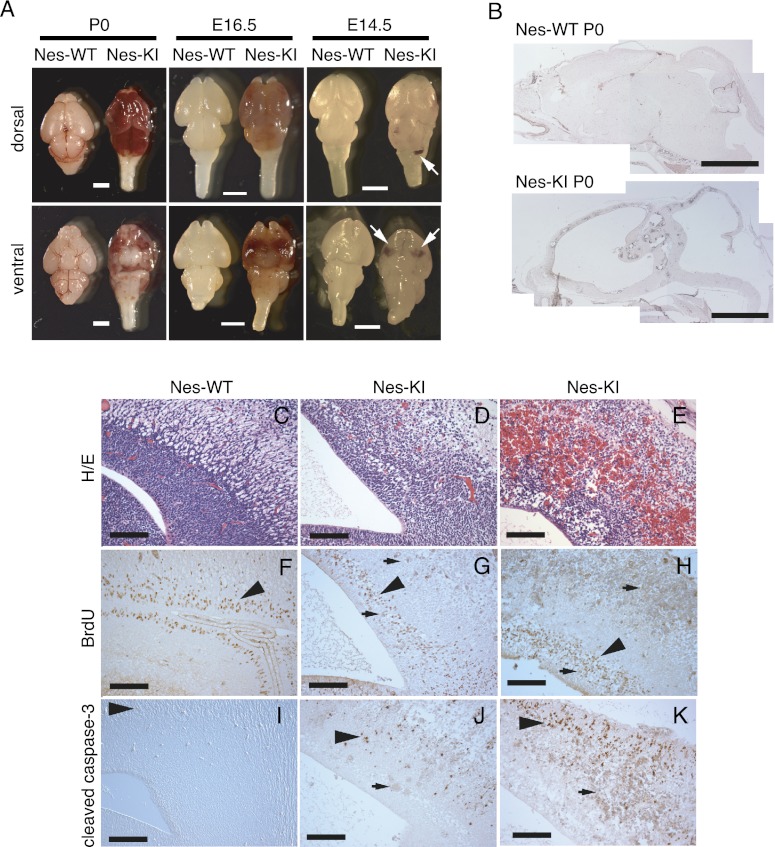
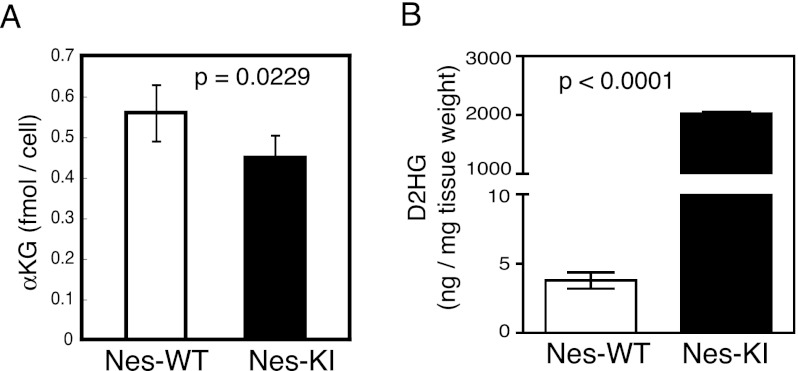
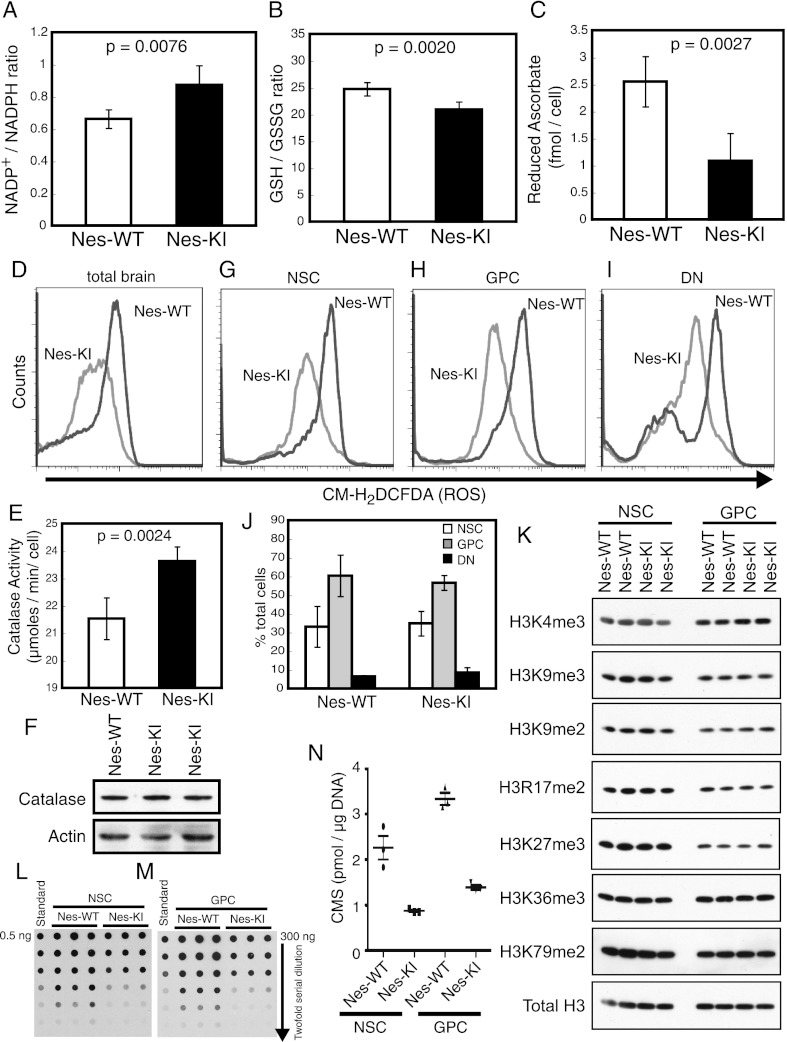
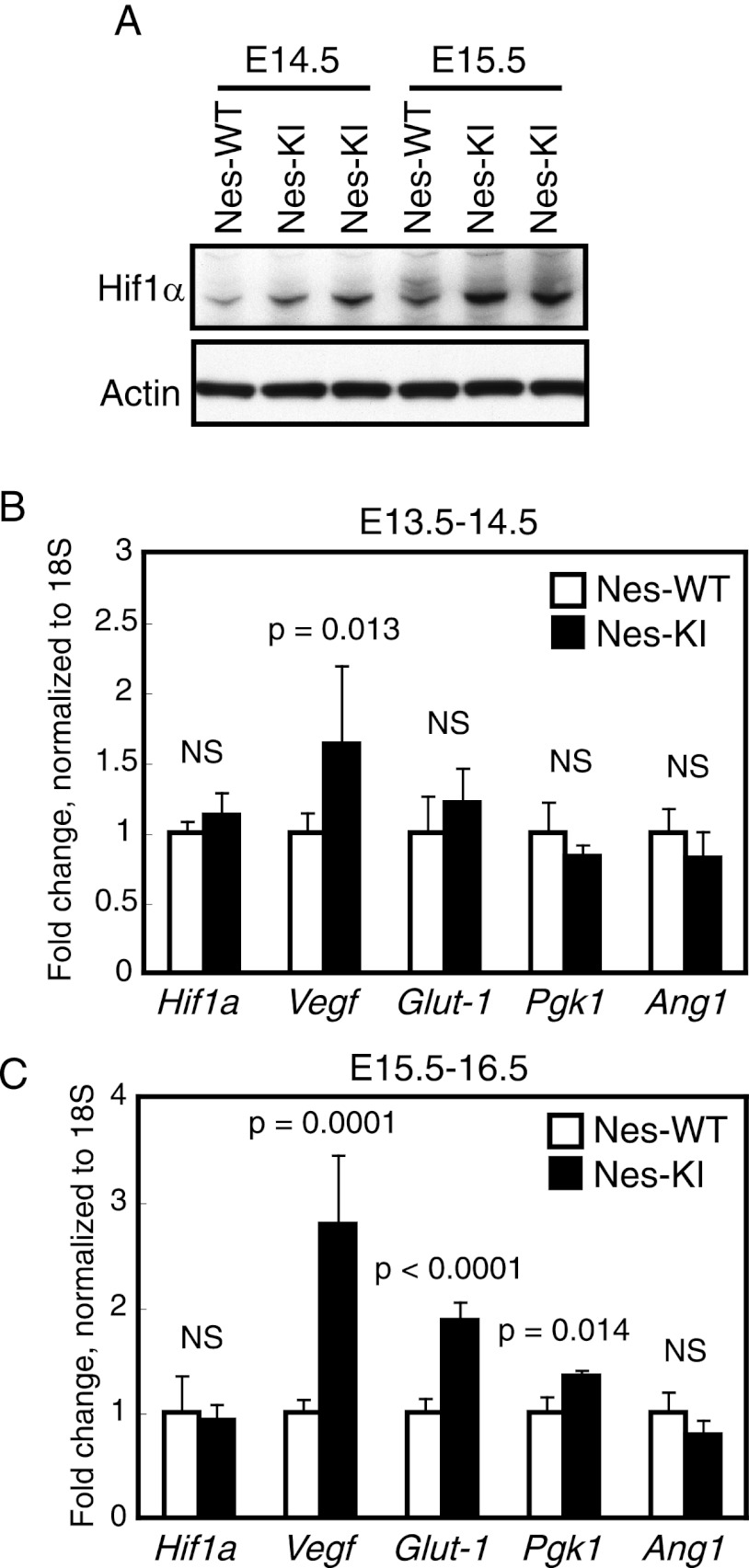
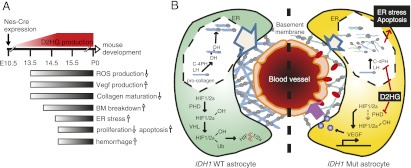
Isocitrate dehydrogenase-1 (IDH1) R132 mutations occur in glioma, but their physiological significance is unknown. Here we describe the generation and characterization of brain-specific Idh1 R132H conditional knock-in (KI) mice. Idh1 mutation results in hemorrhage and perinatal lethality. Surprisingly, intracellular reactive oxygen species (ROS) are attenuated in Idh1-KI brain cells despite an apparent increase in the NADP(+)/NADPH ratio. Idh1-KI cells also show high levels of D-2-hydroxyglutarate (D2HG) that are associated with inhibited prolyl-hydroxylation of hypoxia-inducible transcription factor-1α (Hif1α) and up-regulated Hif1α target gene transcription. Intriguingly, D2HG also blocks prolyl-hydroxylation of collagen, causing a defect in collagen protein maturation. An endoplasmic reticulum (ER) stress response induced by the accumulation of immature collagens may account for the embryonic lethality of these mutants. Importantly, D2HG-mediated impairment of collagen maturation also led to basement membrane (BM) aberrations that could play a part in glioma progression. Our study presents strong in vivo evidence that the D2HG produced by the mutant Idh1 enzyme is responsible for the above effects.
Figures

Similar articles
-
Oncogenic R132 IDH1 Mutations Limit NADPH for De Novo Lipogenesis through (D)2-Hydroxyglutarate Production in Fibrosarcoma Sells.Cell Rep. 2018 Oct 23;25(4):1018-1026.e4. doi: 10.1016/j.celrep.2018.09.074. Cell Rep. 2018. PMID: 30355481 Free PMC article.
-
2-Hydroxyglutarate-Mediated Autophagy of the Endoplasmic Reticulum Leads to an Unusual Downregulation of Phospholipid Biosynthesis in Mutant IDH1 Gliomas.Cancer Res. 2018 May 1;78(9):2290-2304. doi: 10.1158/0008-5472.CAN-17-2926. Epub 2018 Jan 22. Cancer Res. 2018. PMID: 29358170 Free PMC article.
-
Cancer-associated isocitrate dehydrogenase 1 (IDH1) R132H mutation and d-2-hydroxyglutarate stimulate glutamine metabolism under hypoxia.J Biol Chem. 2014 Aug 22;289(34):23318-28. doi: 10.1074/jbc.M114.575183. Epub 2014 Jul 1. J Biol Chem. 2014. PMID: 24986863 Free PMC article.
-
Modeling the diffusion of D-2-hydroxyglutarate from IDH1 mutant gliomas in the central nervous system.Neuro Oncol. 2018 Aug 2;20(9):1197-1206. doi: 10.1093/neuonc/noy051. Neuro Oncol. 2018. PMID: 29660019 Free PMC article.
-
Oncogenic isocitrate dehydrogenase mutations: mechanisms, models, and clinical opportunities.Cancer Discov. 2013 Jul;3(7):730-41. doi: 10.1158/2159-8290.CD-13-0083. Epub 2013 Jun 24. Cancer Discov. 2013. PMID: 23796461 Review.
Cited by
-
The Roles of 2-Hydroxyglutarate.Front Cell Dev Biol. 2021 Mar 26;9:651317. doi: 10.3389/fcell.2021.651317. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33842477 Free PMC article. Review.
-
IDH1 Mutation Induces Reprogramming of Pyruvate Metabolism.Cancer Res. 2015 Aug 1;75(15):2999-3009. doi: 10.1158/0008-5472.CAN-15-0840. Epub 2015 Jun 4. Cancer Res. 2015. PMID: 26045167 Free PMC article.
-
2-Hydroxyglutarate in Cancer Cells.Antioxid Redox Signal. 2020 Nov 1;33(13):903-926. doi: 10.1089/ars.2019.7902. Epub 2020 Jan 22. Antioxid Redox Signal. 2020. PMID: 31847543 Free PMC article. Review.
-
Driver mutations of cancer epigenomes.Protein Cell. 2014 Apr;5(4):265-96. doi: 10.1007/s13238-014-0031-6. Epub 2014 Mar 14. Protein Cell. 2014. PMID: 24622842 Free PMC article.
-
Altered metabolite levels in cancer: implications for tumour biology and cancer therapy.Nat Rev Cancer. 2016 Nov;16(11):680-693. doi: 10.1038/nrc.2016.85. Epub 2016 Sep 23. Nat Rev Cancer. 2016. PMID: 27658530 Review.
References
-
- Amary MF, Bacsi K, Maggiani F, Damato S, Halai D, Berisha F, Pollock R, O'Donnell P, Grigoriadis A, Diss T, et al. 2011a. IDH1 and IDH2 mutations are frequent events in central chondrosarcoma and central and periosteal chondromas but not in other mesenchymal tumours. J Pathol 224: 334–343 - PubMed
-
- Amary MF, Damato S, Halai D, Eskandarpour M, Berisha F, Bonar F, McCarthy S, Fantin VR, Straley KS, Lobo S, et al. 2011b. Ollier disease and Maffucci syndrome are caused by somatic mosaic mutations of IDH1 and IDH2. Nat Genet 43: 1262–1265 - PubMed
-
- Ballabh P, Braun A, Nedergaard M 2004. The blood-brain barrier: An overview: Structure, regulation, and clinical implications. Neurobiol Dis 16: 1–13 - PubMed
-
- Belteki G, Haigh J, Kabacs N, Haigh K, Sison K, Costantini F, Whitsett J, Quaggin SE, Nagy A 2005. Conditional and inducible transgene expression in mice through the combinatorial use of Cre-mediated recombination and tetracycline induction. Nucleic Acids Res 33: e51 doi: 10.1093/nar/gki559 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous