

The Notch signalling system: recent insights into the complexity of a conserved pathway
- PMID: 22868267
- PMCID: PMC4369923
- DOI: 10.1038/nrg3272
The Notch signalling system: recent insights into the complexity of a conserved pathway
Abstract
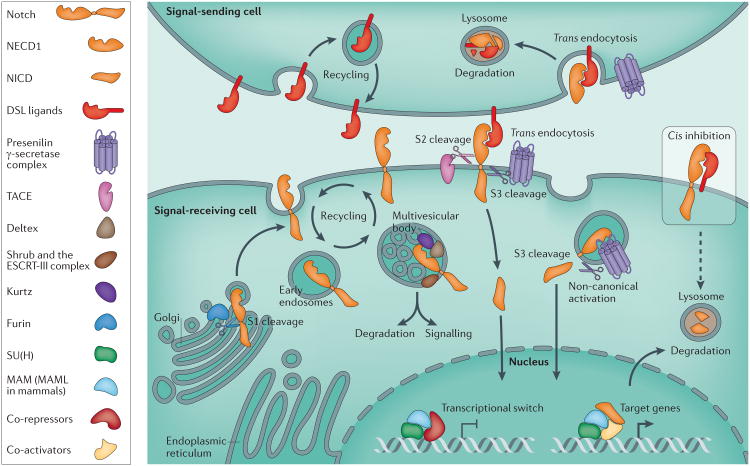
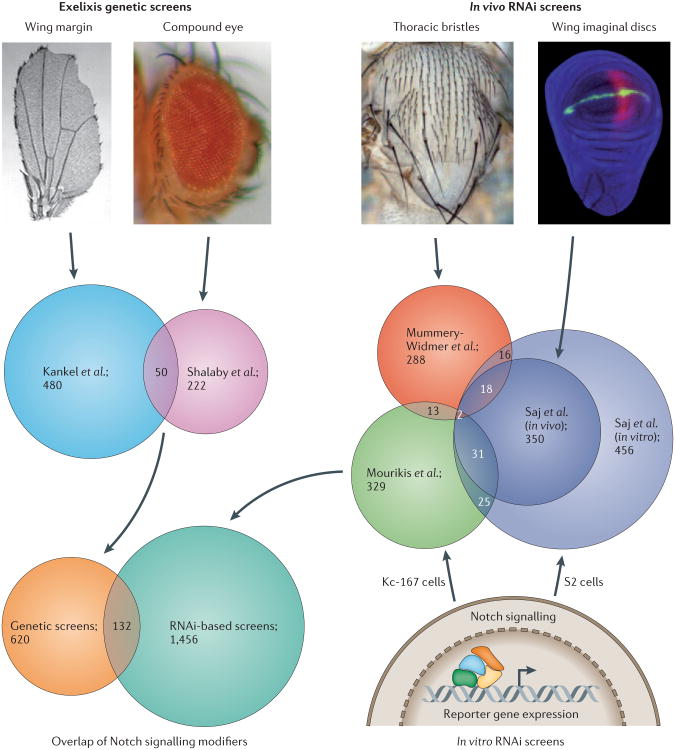
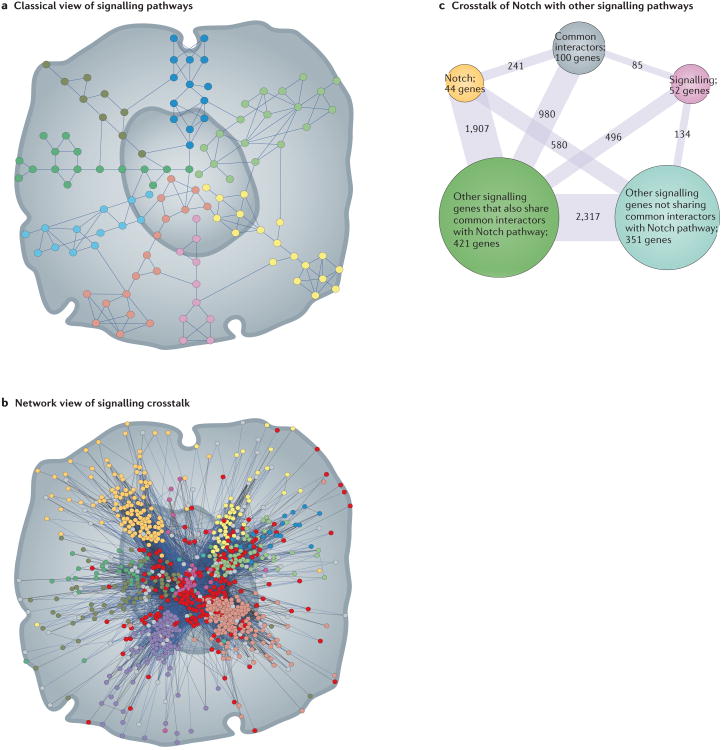
Notch signalling links the fate of one cell to that of an immediate neighbour and consequently controls differentiation, proliferation and apoptotic events in multiple metazoan tissues. Perturbations in this pathway activity have been linked to several human genetic disorders and cancers. Recent genome-scale studies in Drosophila melanogaster have revealed an extraordinarily complex network of genes that can affect Notch activity. This highly interconnected network contrasts our traditional view of the Notch pathway as a simple linear sequence of events. Although we now have an unprecedented insight into the way in which such a fundamental signalling mechanism is controlled by the genome, we are faced with serious challenges in analysing the underlying molecular mechanisms of Notch signal control.
Figures

Similar articles
-
Human Diseases Associated with Notch Signalling: Lessons from Drosophila melanogaster.Front Biosci (Landmark Ed). 2024 Jun 25;29(6):234. doi: 10.31083/j.fbl2906234. Front Biosci (Landmark Ed). 2024. PMID: 38940046 Review.
-
RBPJ/CBF1 interacts with L3MBTL3/MBT1 to promote repression of Notch signaling via histone demethylase KDM1A/LSD1.EMBO J. 2017 Nov 2;36(21):3232-3249. doi: 10.15252/embj.201796525. Epub 2017 Oct 13. EMBO J. 2017. PMID: 29030483 Free PMC article.
-
A targeted in vivo RNAi screen reveals deubiquitinases as new regulators of Notch signaling.G3 (Bethesda). 2012 Dec;2(12):1563-75. doi: 10.1534/g3.112.003780. Epub 2012 Dec 1. G3 (Bethesda). 2012. PMID: 23275879 Free PMC article.
-
Notch signaling: genetics and structure.WormBook. 2013 Jan 17:1-28. doi: 10.1895/wormbook.1.10.2. WormBook. 2013. PMID: 23355521 Free PMC article. Review.
-
The Five Faces of Notch Signalling During Drosophila melanogaster Embryonic CNS Development.Adv Exp Med Biol. 2020;1218:39-58. doi: 10.1007/978-3-030-34436-8_3. Adv Exp Med Biol. 2020. PMID: 32060870 Review.
Cited by
-
Characterization of the Notch pathway in nasal polyps of patients with chronic rhinosinusitis: A pilot study.Physiol Rep. 2022 Aug;10(16):e15403. doi: 10.14814/phy2.15403. Physiol Rep. 2022. PMID: 36029197 Free PMC article.
-
RNA-binding protein Musashi2 induced by RANKL is critical for osteoclast survival.Cell Death Dis. 2016 Jul 21;7(7):e2300. doi: 10.1038/cddis.2016.213. Cell Death Dis. 2016. PMID: 27441652 Free PMC article.
-
Stem cell and progenitor fate in the mammalian intestine: Notch and lateral inhibition in homeostasis and disease.EMBO Rep. 2015 May;16(5):571-81. doi: 10.15252/embr.201540188. Epub 2015 Apr 8. EMBO Rep. 2015. PMID: 25855643 Free PMC article. Review.
-
The ATPase TER94 regulates Notch signaling during Drosophila wing development.Biol Open. 2019 Jan 10;8(1):bio038984. doi: 10.1242/bio.038984. Biol Open. 2019. PMID: 30530809 Free PMC article.
-
RBM8A Promotes Glioblastoma Growth and Invasion Through the Notch/STAT3 Pathway.Front Oncol. 2021 Nov 4;11:736941. doi: 10.3389/fonc.2021.736941. eCollection 2021. Front Oncol. 2021. PMID: 34804926 Free PMC article.
References
-
- Dexter JS. The analysis of a case of continuous variation in Drosophila by a study of its linkage relations. Am Nat. 1914;48:712–758.
-
- Morgan TH, Bridges CB. Sex-Linked Inheritance in Drosophila. Carnegie Institute of Washington; 1916.
-
- Artavanis-Tsakonas S, Muskavitch MA. Notch: the past, the present, and the future. Curr Top Dev Biol. 2010;92:1–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
