

Identification of regulators of polyploidization presents therapeutic targets for treatment of AMKL
- PMID: 22863010
- PMCID: PMC3613864
- DOI: 10.1016/j.cell.2012.06.032
Identification of regulators of polyploidization presents therapeutic targets for treatment of AMKL
Abstract
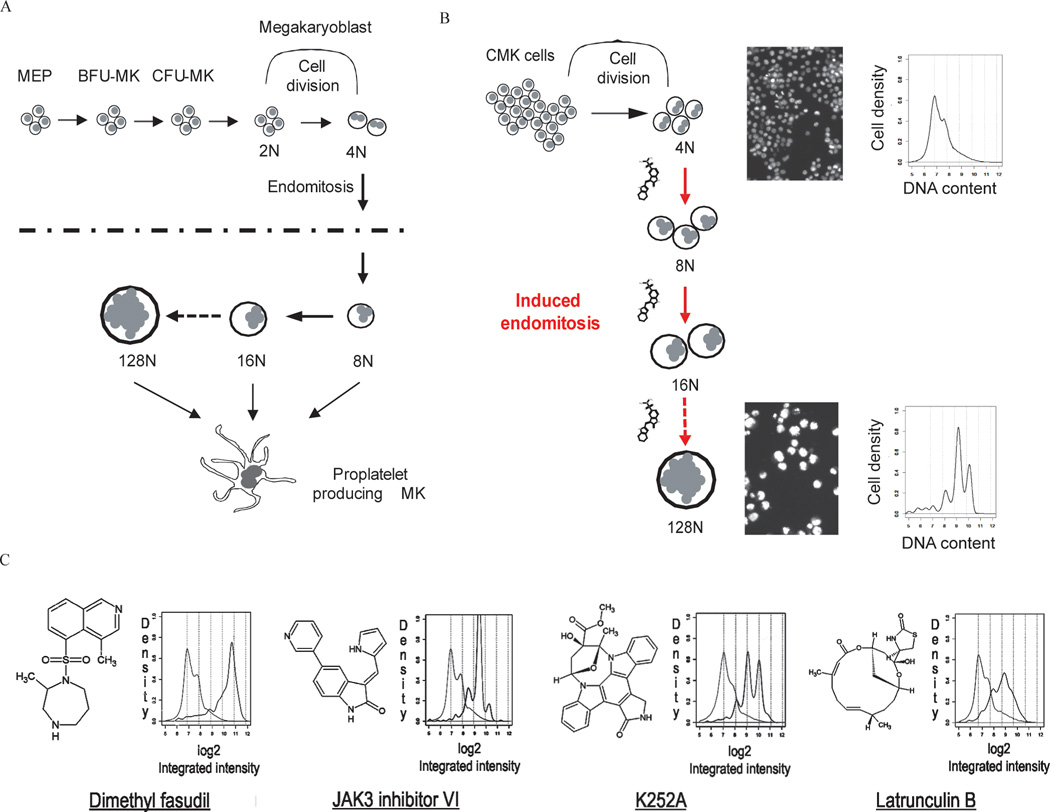
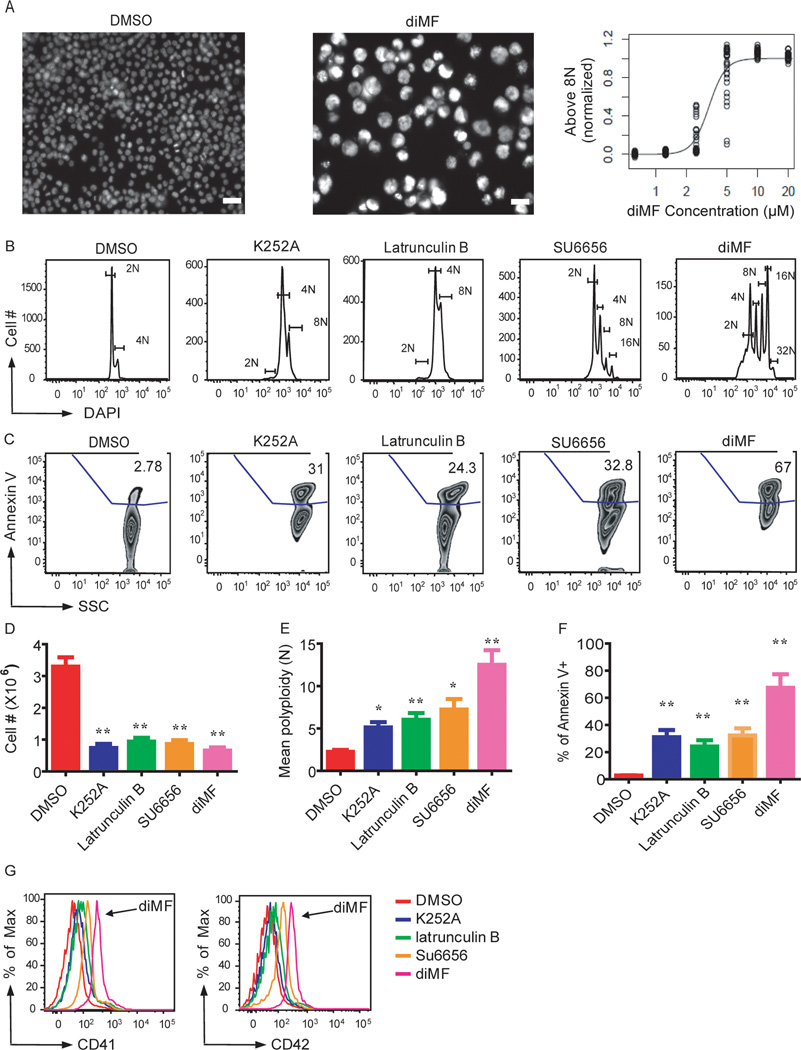
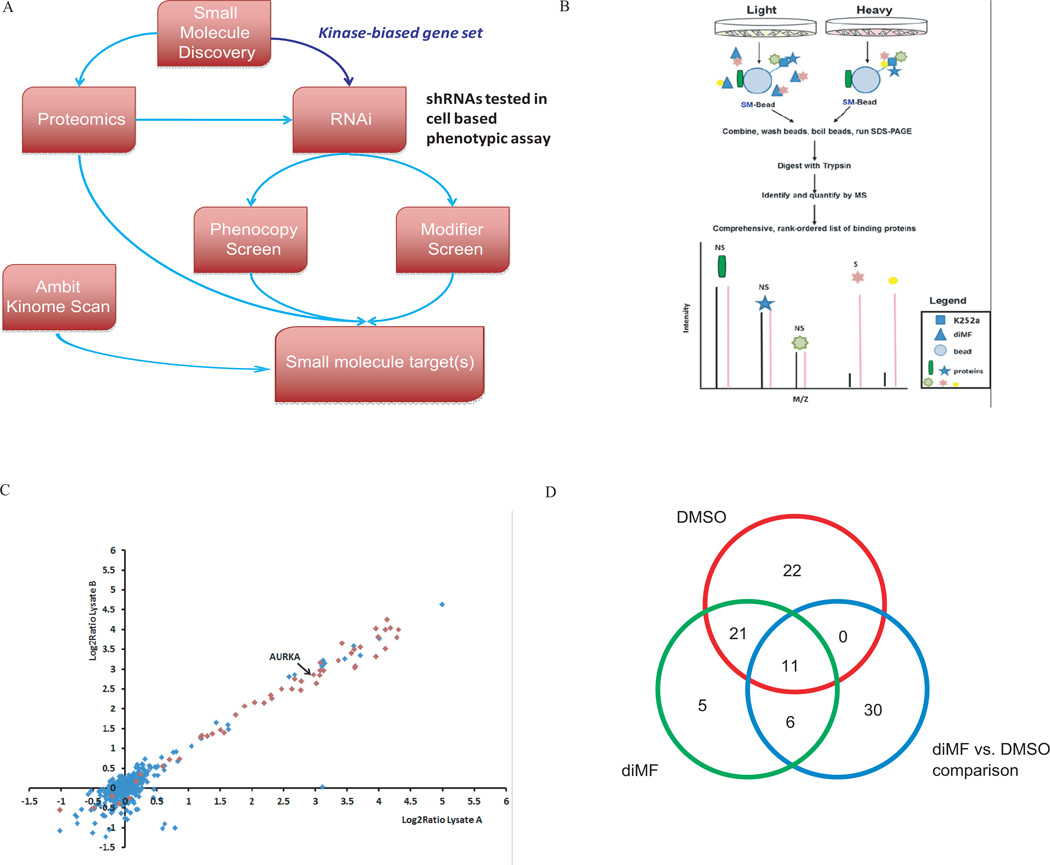
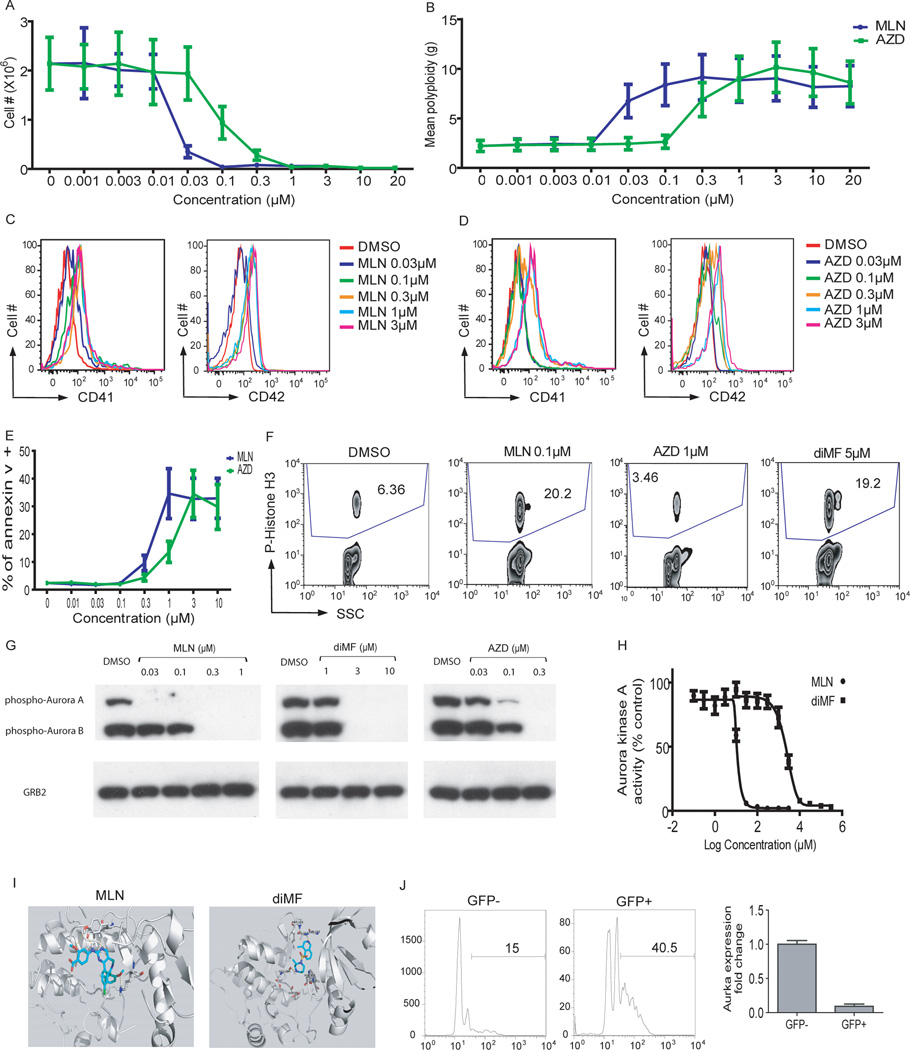
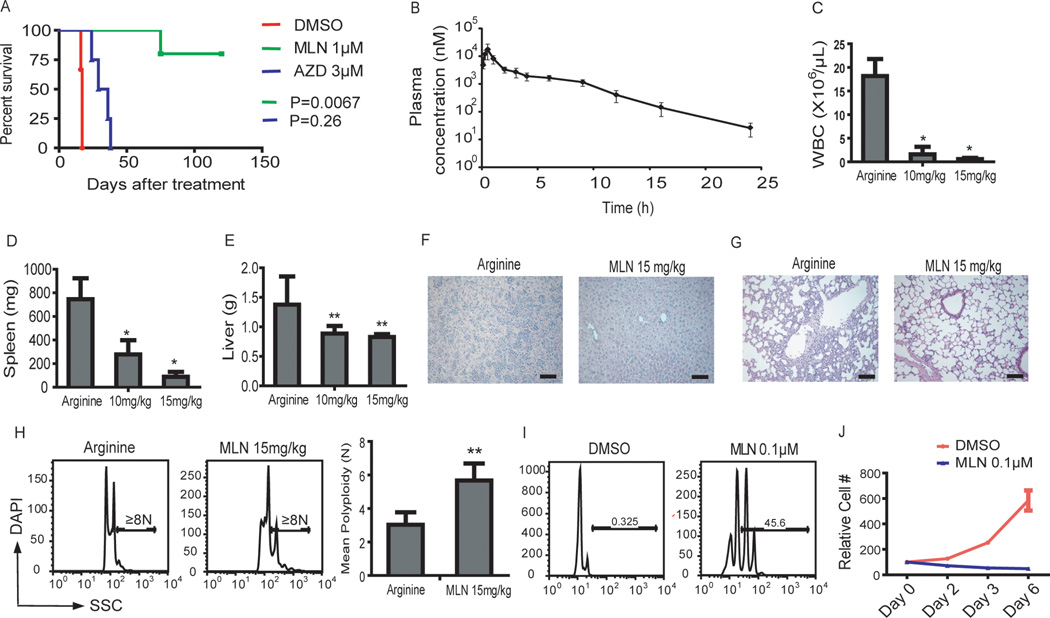
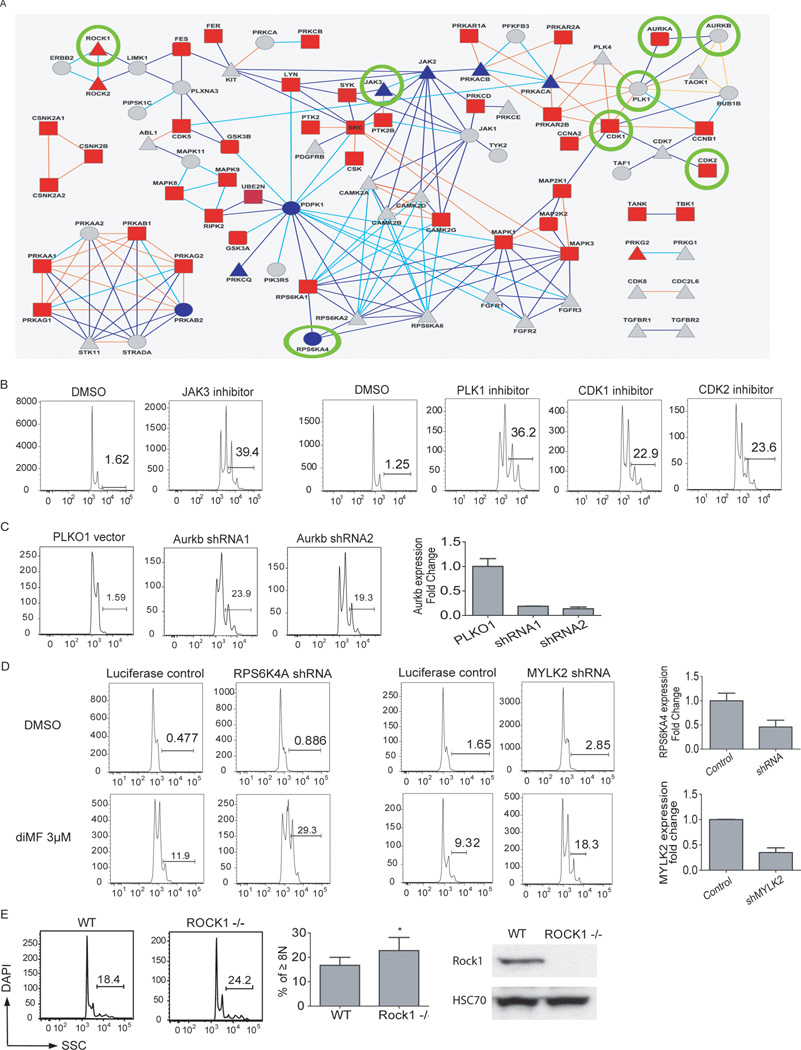
The mechanism by which cells decide to skip mitosis to become polyploid is largely undefined. Here we used a high-content image-based screen to identify small-molecule probes that induce polyploidization of megakaryocytic leukemia cells and serve as perturbagens to help understand this process. Our study implicates five networks of kinases that regulate the switch to polyploidy. Moreover, we find that dimethylfasudil (diMF, H-1152P) selectively increased polyploidization, mature cell-surface marker expression, and apoptosis of malignant megakaryocytes. An integrated target identification approach employing proteomic and shRNA screening revealed that a major target of diMF is Aurora kinase A (AURKA). We further find that MLN8237 (Alisertib), a selective inhibitor of AURKA, induced polyploidization and expression of mature megakaryocyte markers in acute megakaryocytic leukemia (AMKL) blasts and displayed potent anti-AMKL activity in vivo. Our findings provide a rationale to support clinical trials of MLN8237 and other inducers of polyploidization and differentiation in AMKL.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures

Similar articles
-
Fasudil promotes polyploidization of megakaryoblasts in an acute megakaryocyte leukemia model.Naunyn Schmiedebergs Arch Pharmacol. 2023 Nov;396(11):3101-3110. doi: 10.1007/s00210-023-02513-6. Epub 2023 May 10. Naunyn Schmiedebergs Arch Pharmacol. 2023. PMID: 37162543
-
Molecular pathways: induction of polyploidy as a novel differentiation therapy for leukemia.Clin Cancer Res. 2013 Nov 15;19(22):6084-8. doi: 10.1158/1078-0432.CCR-12-2604. Epub 2013 Aug 20. Clin Cancer Res. 2013. PMID: 23963861 Free PMC article.
-
Characterization of novel genomic alterations and therapeutic approaches using acute megakaryoblastic leukemia xenograft models.J Exp Med. 2012 Oct 22;209(11):2017-31. doi: 10.1084/jem.20121343. Epub 2012 Oct 8. J Exp Med. 2012. PMID: 23045605 Free PMC article.
-
Pediatric Acute Megakaryoblastic Leukemia: Multitasking Fusion Proteins and Oncogenic Cooperations.Trends Cancer. 2017 Sep;3(9):631-642. doi: 10.1016/j.trecan.2017.07.003. Epub 2017 Aug 18. Trends Cancer. 2017. PMID: 28867167 Review.
-
Acute megakaryocytic leukemia: What have we learned.Blood Rev. 2016 Jan;30(1):49-53. doi: 10.1016/j.blre.2015.07.005. Epub 2015 Jul 18. Blood Rev. 2016. PMID: 26228843 Review.
Cited by
-
Targeting megakaryocytic-induced fibrosis in myeloproliferative neoplasms by AURKA inhibition.Nat Med. 2015 Dec;21(12):1473-80. doi: 10.1038/nm.3995. Epub 2015 Nov 16. Nat Med. 2015. PMID: 26569382 Free PMC article.
-
Fasudil promotes polyploidization of megakaryoblasts in an acute megakaryocyte leukemia model.Naunyn Schmiedebergs Arch Pharmacol. 2023 Nov;396(11):3101-3110. doi: 10.1007/s00210-023-02513-6. Epub 2023 May 10. Naunyn Schmiedebergs Arch Pharmacol. 2023. PMID: 37162543
-
Down syndrome and leukemia: insights into leukemogenesis and translational targets.Transl Pediatr. 2015 Apr;4(2):76-92. doi: 10.3978/j.issn.2224-4336.2015.03.03. Transl Pediatr. 2015. PMID: 26835364 Free PMC article. Review.
-
Heterogeneous cytogenetic subgroups and outcomes in childhood acute megakaryoblastic leukemia: a retrospective international study.Blood. 2015 Sep 24;126(13):1575-84. doi: 10.1182/blood-2015-02-629204. Epub 2015 Jul 27. Blood. 2015. PMID: 26215111 Free PMC article.
-
Molecular pathways: induction of polyploidy as a novel differentiation therapy for leukemia.Clin Cancer Res. 2013 Nov 15;19(22):6084-8. doi: 10.1158/1078-0432.CCR-12-2604. Epub 2013 Aug 20. Clin Cancer Res. 2013. PMID: 23963861 Free PMC article.
References
-
- Bluteau D, Lordier L, Di Stefano A, Chang Y, Raslova H, Debili N, Vainchenker W. Regulation of megakaryocyte maturation and platelet formation. J Thromb Haemost. 2009;7(Suppl 1):227–234. - PubMed
-
- Carmena M, Earnshaw WC. The cellular geography of aurora kinases. Nat Rev Mol Cell Biol. 2003;4:842–854. - PubMed
-
- D'Alise AM, Amabile G, Iovino M, Di Giorgio FP, Bartiromo M, Sessa F, Villa F, Musacchio A, Cortese R. Reversine, a novel Aurora kinases inhibitor, inhibits colony formation of human acute myeloid leukemia cells. Mol Cancer Ther. 2008;7:1140–1149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL075816/HL/NHLBI NIH HHS/United States
- HL077177/HL/NHLBI NIH HHS/United States
- U54 HG005032/HG/NHGRI NIH HHS/United States
- N01--CO-12400/CO/NCI NIH HHS/United States
- CA101774/CA/NCI NIH HHS/United States
- RL1-GM084437/GM/NIGMS NIH HHS/United States
- R01 GM089652/GM/NIGMS NIH HHS/United States
- P30 DK056465/DK/NIDDK NIH HHS/United States
- RL1-CA133834/CA/NCI NIH HHS/United States
- HL075816/HL/NHLBI NIH HHS/United States
- U10 CA98543/CA/NCI NIH HHS/United States
- R01 HL081111/HL/NHLBI NIH HHS/United States
- UL1-DE019585/DE/NIDCR NIH HHS/United States
- R56 HL075816/HL/NHLBI NIH HHS/United States
- R01 CA101774/CA/NCI NIH HHS/United States
- UL1 DE019585/DE/NIDCR NIH HHS/United States
- P01 HL085098/HL/NHLBI NIH HHS/United States
- RL1 GM084437/GM/NIGMS NIH HHS/United States
- RL1-HG004671/HG/NHGRI NIH HHS/United States
- U10 CA098543/CA/NCI NIH HHS/United States
- GM089652/GM/NIGMS NIH HHS/United States
- N01CO12400/CA/NCI NIH HHS/United States
- R01 HL077177/HL/NHLBI NIH HHS/United States
- RL1 HG004671/HG/NHGRI NIH HHS/United States
- RL1 CA133834/CA/NCI NIH HHS/United States
- HL081111/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
