

Programmed elimination of cells by caspase-independent cell extrusion in C. elegans
- PMID: 22801495
- PMCID: PMC3416925
- DOI: 10.1038/nature11240
Programmed elimination of cells by caspase-independent cell extrusion in C. elegans
Abstract
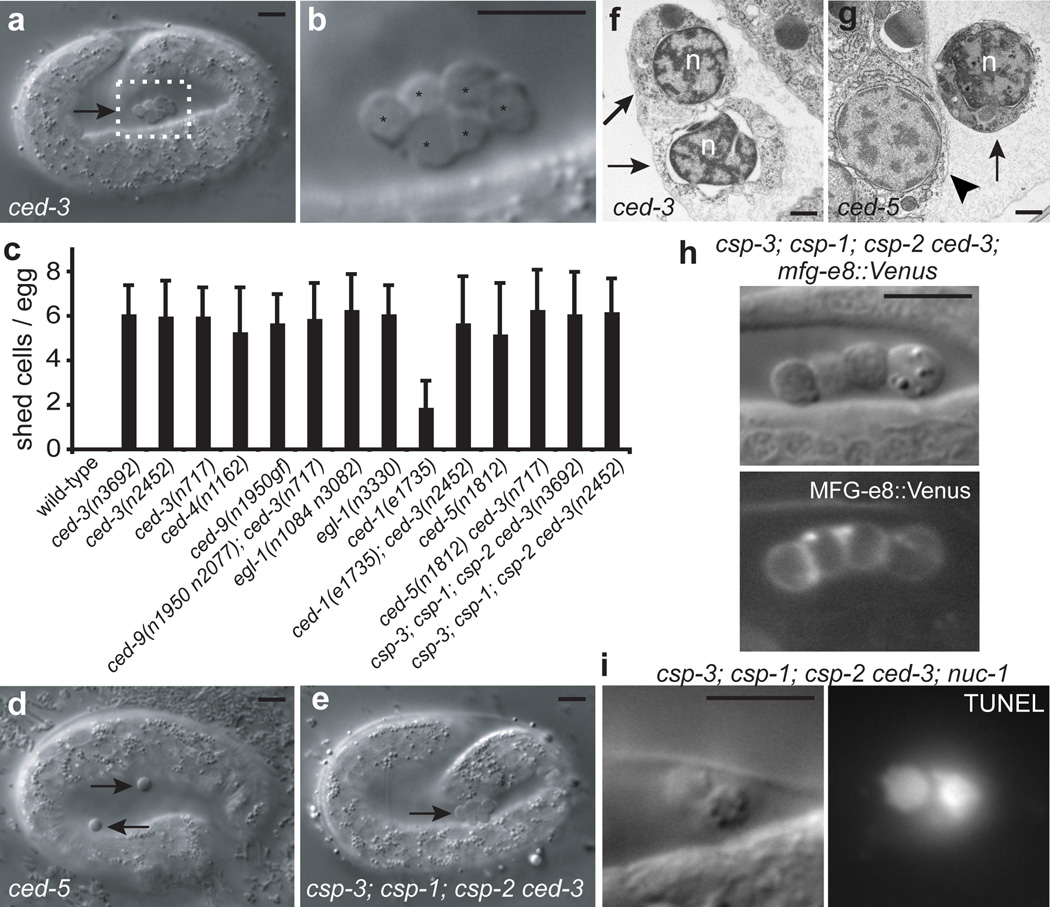
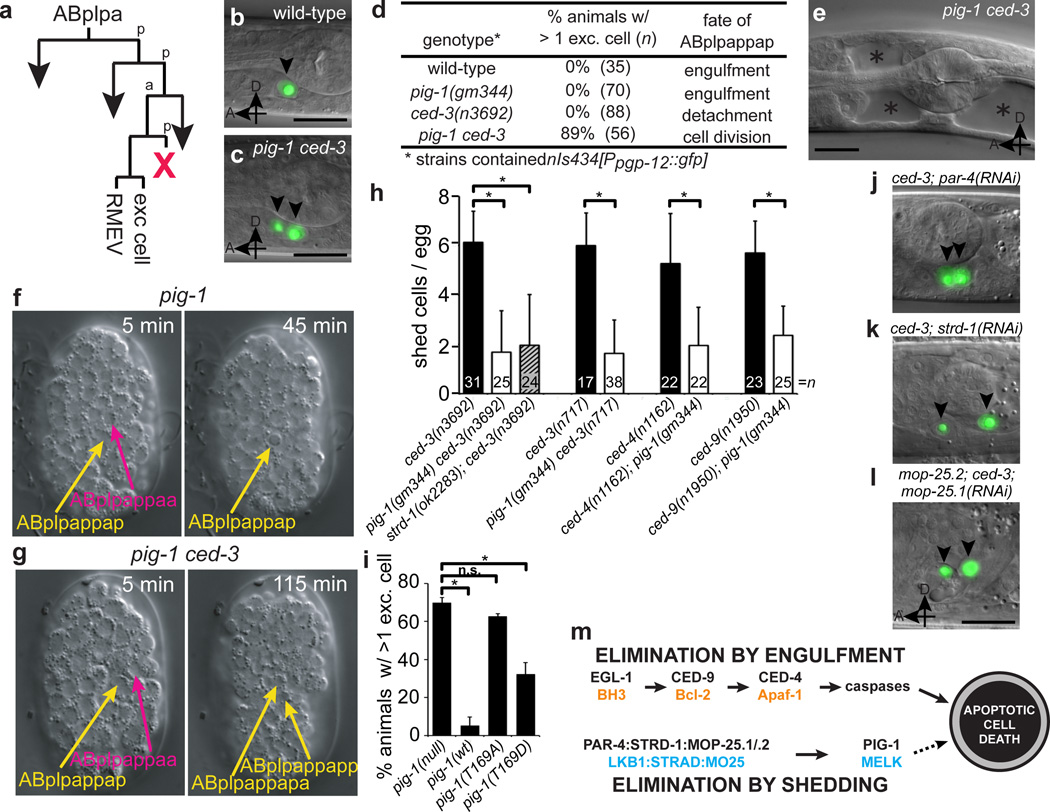
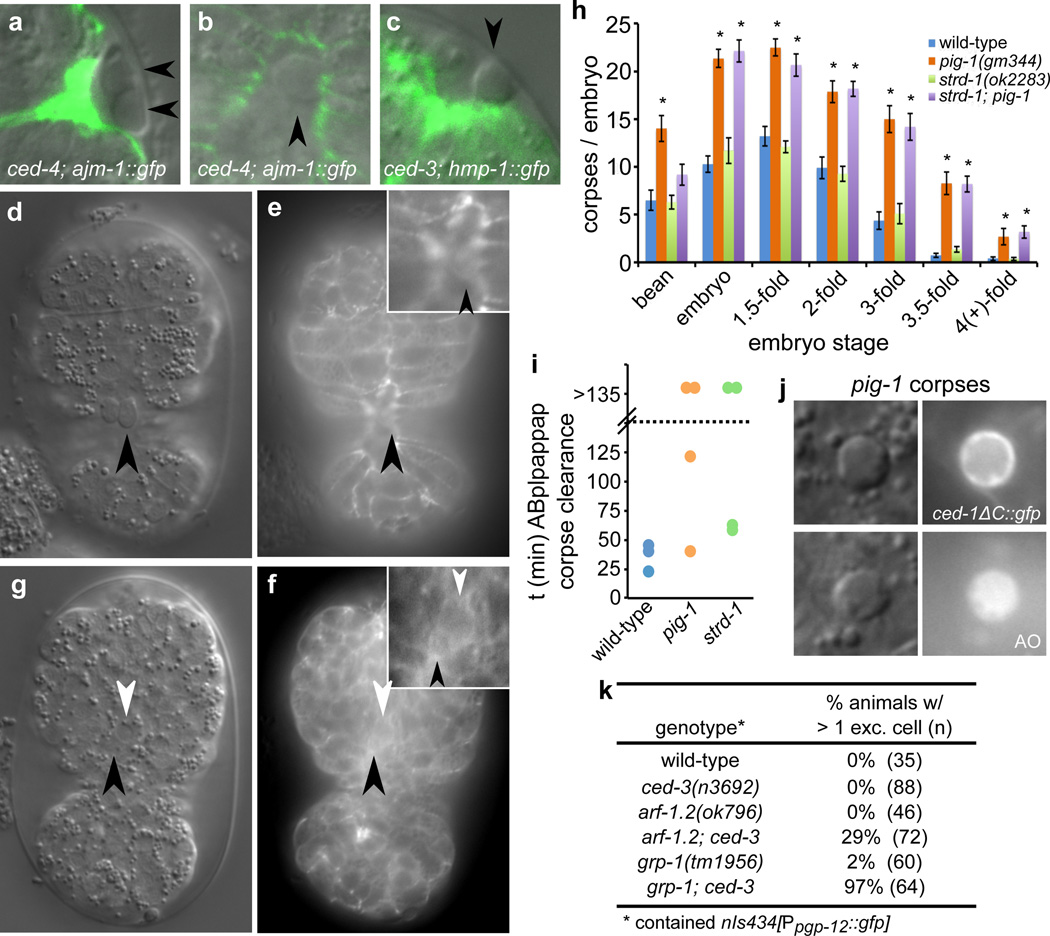
The elimination of unnecessary or defective cells from metazoans occurs during normal development and tissue homeostasis, as well as in response to infection or cellular damage. Although many cells are removed through caspase-mediated apoptosis followed by phagocytosis by engulfing cells, other mechanisms of cell elimination occur, including the extrusion of cells from epithelia through a poorly understood, possibly caspase-independent, process. Here we identify a mechanism of cell extrusion that is caspase independent and that can eliminate a subset of the Caenorhabditis elegans cells programmed to die during embryonic development. In wild-type animals, these cells die soon after their generation through caspase-mediated apoptosis. However, in mutants lacking all four C. elegans caspase genes, these cells are eliminated by being extruded from the developing embryo into the extra-embryonic space of the egg. The shed cells show apoptosis-like cytological and morphological characteristics, indicating that apoptosis can occur in the absence of caspases in C. elegans. We describe a kinase pathway required for cell extrusion involving PAR-4, STRD-1 and MOP-25.1/-25.2, the C. elegans homologues of the mammalian tumour-suppressor kinase LKB1 and its binding partners STRADα and MO25α. The AMPK-related kinase PIG-1, a possible target of the PAR-4–STRD-1–MOP-25 kinase complex, is also required for cell shedding. PIG-1 promotes shed-cell detachment by preventing the cell-surface expression of cell-adhesion molecules. Our findings reveal a mechanism for apoptotic cell elimination that is fundamentally distinct from that of canonical programmed cell death.
Figures

Comment in
-
Programmed cell death: a new way worms get rid of unwanted cells.Curr Biol. 2012 Oct 9;22(19):R844-6. doi: 10.1016/j.cub.2012.08.013. Curr Biol. 2012. PMID: 23058805 Free PMC article.
Similar articles
-
Both the caspase CSP-1 and a caspase-independent pathway promote programmed cell death in parallel to the canonical pathway for apoptosis in Caenorhabditis elegans.PLoS Genet. 2013;9(3):e1003341. doi: 10.1371/journal.pgen.1003341. Epub 2013 Mar 7. PLoS Genet. 2013. PMID: 23505386 Free PMC article.
-
Programmed cell death: a new way worms get rid of unwanted cells.Curr Biol. 2012 Oct 9;22(19):R844-6. doi: 10.1016/j.cub.2012.08.013. Curr Biol. 2012. PMID: 23058805 Free PMC article.
-
Caenorhabditis elegans PIG-1/MELK acts in a conserved PAR-4/LKB1 polarity pathway to promote asymmetric neuroblast divisions.Genetics. 2013 Mar;193(3):897-909. doi: 10.1534/genetics.112.148106. Epub 2012 Dec 24. Genetics. 2013. PMID: 23267054 Free PMC article.
-
Cell death specification in C. elegans.Cell Cycle. 2008 Aug 15;7(16):2479-84. doi: 10.4161/cc.7.16.6479. Epub 2008 Aug 19. Cell Cycle. 2008. PMID: 18719375 Free PMC article. Review.
-
2:1 Stoichiometry of the CED-4-CED-9 complex and the tetrameric CED-4: insights into the regulation of CED-3 activation.Cell Cycle. 2006 Jan;5(1):31-4. doi: 10.4161/cc.5.1.2263. Epub 2006 Jan 18. Cell Cycle. 2006. PMID: 16294007 Review.
Cited by
-
The Par3-like polarity protein Par3L is essential for mammary stem cell maintenance.Nat Cell Biol. 2014 Jun;16(6):529-37. doi: 10.1038/ncb2969. Epub 2014 May 25. Nat Cell Biol. 2014. PMID: 24859006 Free PMC article.
-
Cloning, tissue distribution, expression pattern, and function of porcine maternal embryonic leucine zipper kinase.Ann Transl Med. 2020 Mar;8(5):239. doi: 10.21037/atm.2020.03.46. Ann Transl Med. 2020. PMID: 32309386 Free PMC article.
-
Programmed cell death and clearance of cell corpses in Caenorhabditis elegans.Cell Mol Life Sci. 2016 Jun;73(11-12):2221-36. doi: 10.1007/s00018-016-2196-z. Epub 2016 Apr 5. Cell Mol Life Sci. 2016. PMID: 27048817 Free PMC article. Review.
-
Programmed cell removal by calreticulin in tissue homeostasis and cancer.Nat Commun. 2018 Aug 10;9(1):3194. doi: 10.1038/s41467-018-05211-7. Nat Commun. 2018. PMID: 30097573 Free PMC article.
-
Autophagy mediates phosphatidylserine exposure and phagosome degradation during apoptosis through specific functions of GABARAP/LGG-1 and LC3/LGG-2.Autophagy. 2019 Feb;15(2):228-241. doi: 10.1080/15548627.2018.1512452. Epub 2018 Sep 10. Autophagy. 2019. PMID: 30160610 Free PMC article.
References
-
- Reddien PW, Horvitz HR. The engulfment process of programmed cell death in Caenorhabditis elegans. Annu. Rev. Cell Dev. Biol. 2004;20:193–221. - PubMed
-
- Rosenblatt J, Raff MC, Cramer LP. An epithelial cell destined for apoptosis signals its neighbors to extrude it by an actin- and myosin-dependent mechanism. Curr. Biol. 2001;11:1847–1857. - PubMed
-
- Ellis HM, Horvitz HR. Genetic control of programmed cell death in the nematode C. elegans. Cell. 1986;44:817–829. - PubMed
Methods References
-
- Schwartz HT. A protocol describing pharynx counts and a review of other assays of apoptotic cell death in the nematode worm Caenorhabditis elegans. Nat Protoc. 2007;2:705–714. - PubMed
-
- Fraser AG, et al. Functional genomic analysis of C. elegans chromosome I by systematic RNA interference. Nature. 2000;408:325–330. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
