

Commitment to glycolysis sustains survival of NO-producing inflammatory dendritic cells
- PMID: 22786879
- PMCID: PMC3423780
- DOI: 10.1182/blood-2012-03-419747
Commitment to glycolysis sustains survival of NO-producing inflammatory dendritic cells
Abstract
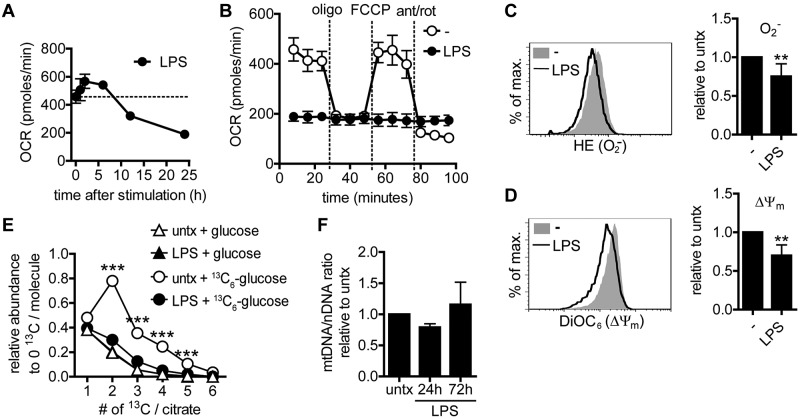
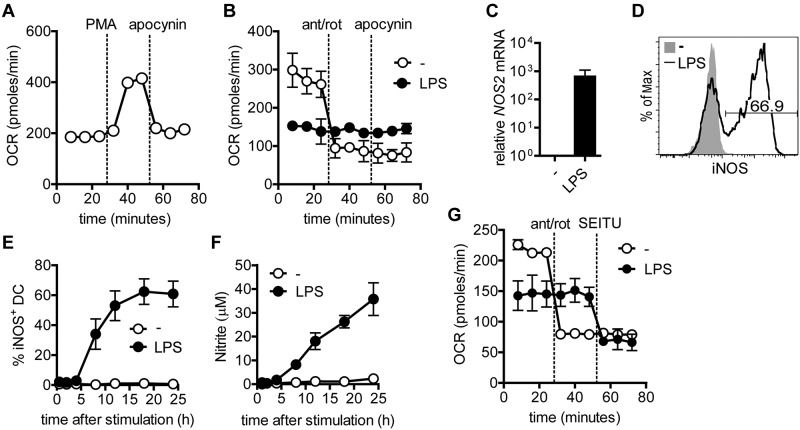
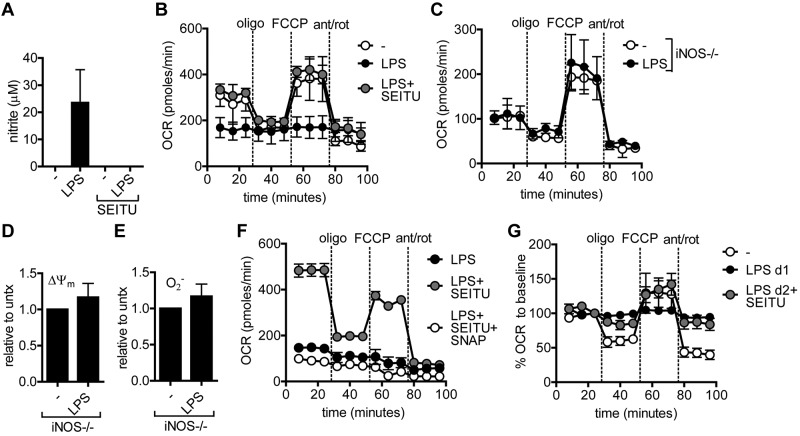
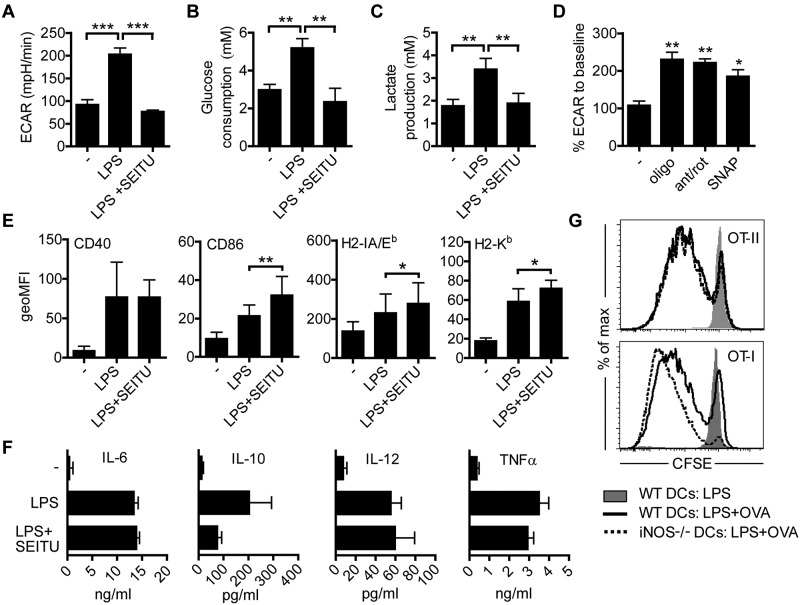
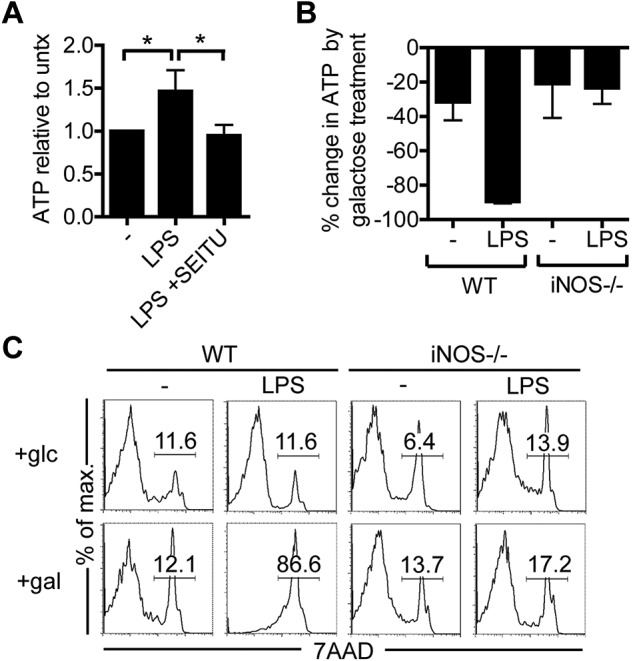
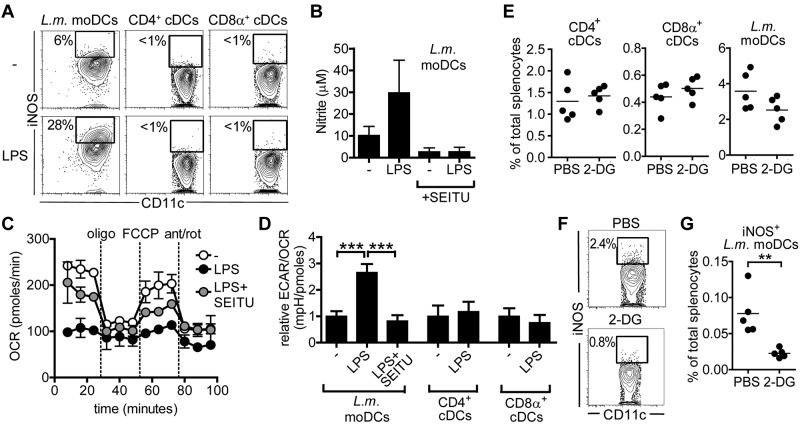
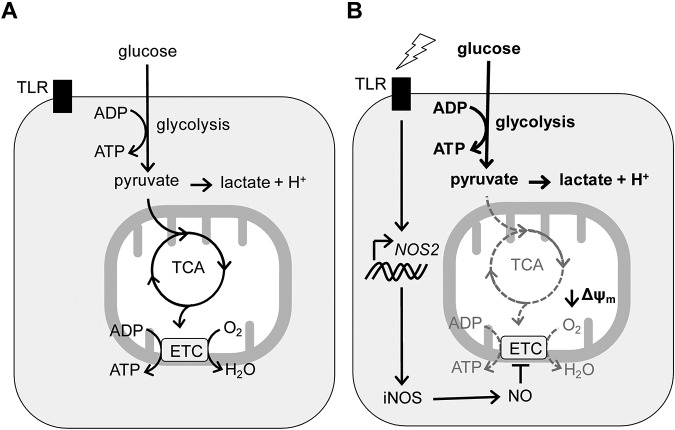
TLR agonists initiate a rapid activation program in dendritic cells (DCs) that requires support from metabolic and bioenergetic resources. We found previously that TLR signaling promotes aerobic glycolysis and a decline in oxidative phosphorylation (OXHPOS) and that glucose restriction prevents activation and leads to premature cell death. However, it remained unclear why the decrease in OXPHOS occurs under these circumstances. Using real-time metabolic flux analysis, in the present study, we show that mitochondrial activity is lost progressively after activation by TLR agonists in inflammatory blood monocyte-derived DCs that express inducible NO synthase. We found that this is because of inhibition of OXPHOS by NO and that the switch to glycolysis is a survival response that serves to maintain ATP levels when OXPHOS is inhibited. Our data identify NO as a profound metabolic regulator in inflammatory monocyte-derived DCs.
Figures
Similar articles
-
Mechanistic target of rapamycin inhibition extends cellular lifespan in dendritic cells by preserving mitochondrial function.J Immunol. 2014 Sep 15;193(6):2821-30. doi: 10.4049/jimmunol.1302498. Epub 2014 Aug 8. J Immunol. 2014. PMID: 25108022 Free PMC article.
-
Human Plasmacytoid and Monocyte-Derived Dendritic Cells Display Distinct Metabolic Profile Upon RIG-I Activation.Front Immunol. 2018 Dec 21;9:3070. doi: 10.3389/fimmu.2018.03070. eCollection 2018. Front Immunol. 2018. PMID: 30622542 Free PMC article.
-
Divergent Genetic Regulation of Nitric Oxide Production between C57BL/6J and Wild-Derived PWD/PhJ Mice Controls Postactivation Mitochondrial Metabolism, Cell Survival, and Bacterial Resistance in Dendritic Cells.J Immunol. 2022 Jan 1;208(1):97-109. doi: 10.4049/jimmunol.2100375. Epub 2021 Dec 6. J Immunol. 2022. PMID: 34872978 Free PMC article.
-
The role of nitric oxide in metabolic regulation of Dendritic cell immune function.Cancer Lett. 2018 Jan 1;412:236-242. doi: 10.1016/j.canlet.2017.10.032. Epub 2017 Oct 26. Cancer Lett. 2018. PMID: 29107106 Free PMC article. Review.
-
Differentiation and function of mouse monocyte-derived dendritic cells in steady state and inflammation.Immunol Rev. 2010 Mar;234(1):90-104. doi: 10.1111/j.0105-2896.2009.00876.x. Immunol Rev. 2010. PMID: 20193014 Review.
Cited by
-
Dendritic cell metabolism.Nat Rev Immunol. 2015 Jan;15(1):18-29. doi: 10.1038/nri3771. Nat Rev Immunol. 2015. PMID: 25534620 Free PMC article. Review.
-
Comparative evaluation of itaconate and its derivatives reveals divergent inflammasome and type I interferon regulation in macrophages.Nat Metab. 2020 Jul;2(7):594-602. doi: 10.1038/s42255-020-0210-0. Epub 2020 May 25. Nat Metab. 2020. PMID: 32694786 Free PMC article.
-
Mitochondria as Key Players in the Pathogenesis and Treatment of Rheumatoid Arthritis.Front Immunol. 2021 Apr 29;12:673916. doi: 10.3389/fimmu.2021.673916. eCollection 2021. Front Immunol. 2021. PMID: 33995417 Free PMC article. Review.
-
The Evolving Role of Dendritic Cells in Atherosclerosis.Int J Mol Sci. 2024 Feb 19;25(4):2450. doi: 10.3390/ijms25042450. Int J Mol Sci. 2024. PMID: 38397127 Free PMC article. Review.
-
Succinate Dehydrogenase Supports Metabolic Repurposing of Mitochondria to Drive Inflammatory Macrophages.Cell. 2016 Oct 6;167(2):457-470.e13. doi: 10.1016/j.cell.2016.08.064. Epub 2016 Sep 22. Cell. 2016. PMID: 27667687 Free PMC article.
References
-
- Kapsenberg ML. Dendritic-cell control of pathogen-driven T-cell polarization. Nat Rev Immunol. 2003;3(12):984–993. - PubMed
-
- Joffre O, Nolte MA, Sporri R, Reis e Sousa C. Inflammatory signals in dendritic cell activation and the induction of adaptive immunity. Immunol Rev. 2009;227(1):234–247. - PubMed
-
- Amati L, Pepe M, Passeri ME, Mastronardi ML, Jirillo E, Covelli V. Toll-like receptor signaling mechanisms involved in dendritic cell activation: potential therapeutic control of T cell polarization. Curr Pharm Des. 2006;12(32):4247–4254. - PubMed
-
- Warburg O. On the origin of cancer cells. Science. 1956;123(3191):309–314. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
