

Control of vertebrate intraflagellar transport by the planar cell polarity effector Fuz
- PMID: 22778277
- PMCID: PMC3392940
- DOI: 10.1083/jcb.201204072
Control of vertebrate intraflagellar transport by the planar cell polarity effector Fuz
Abstract
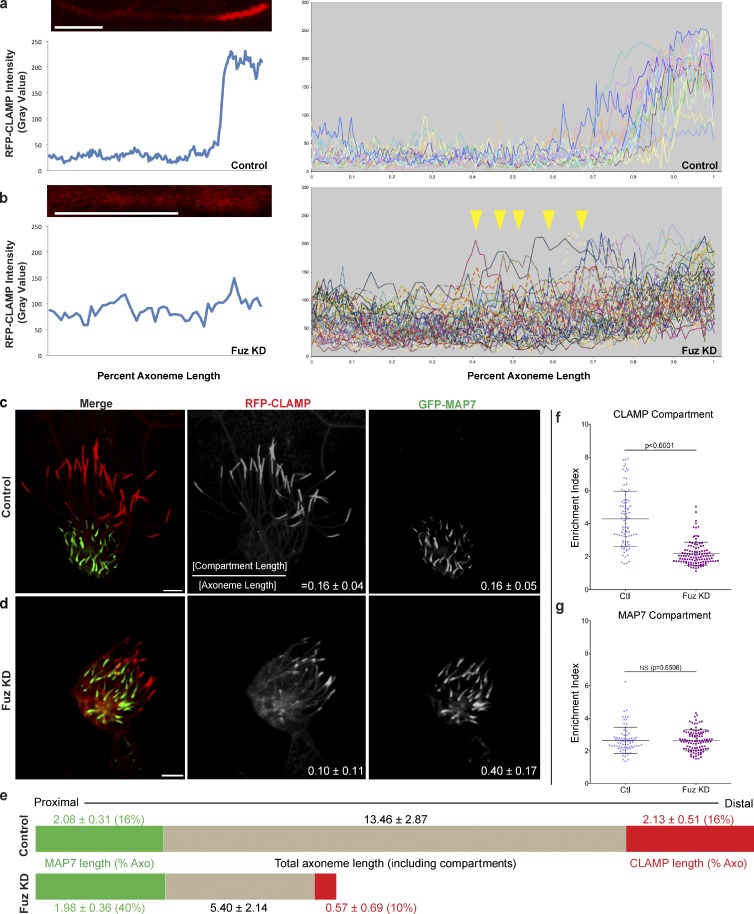
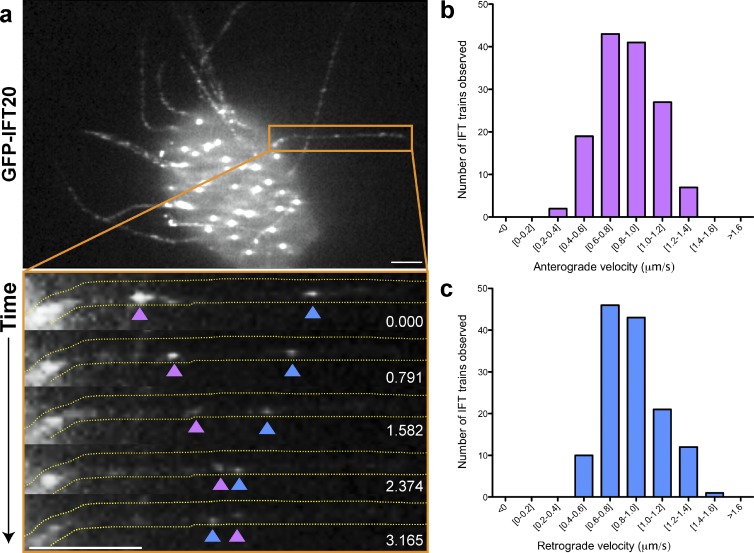
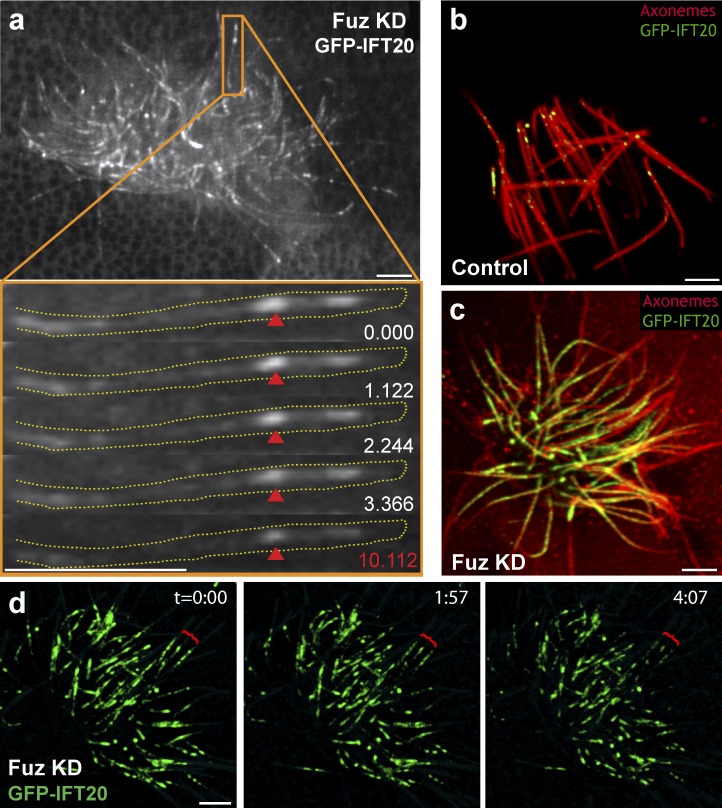
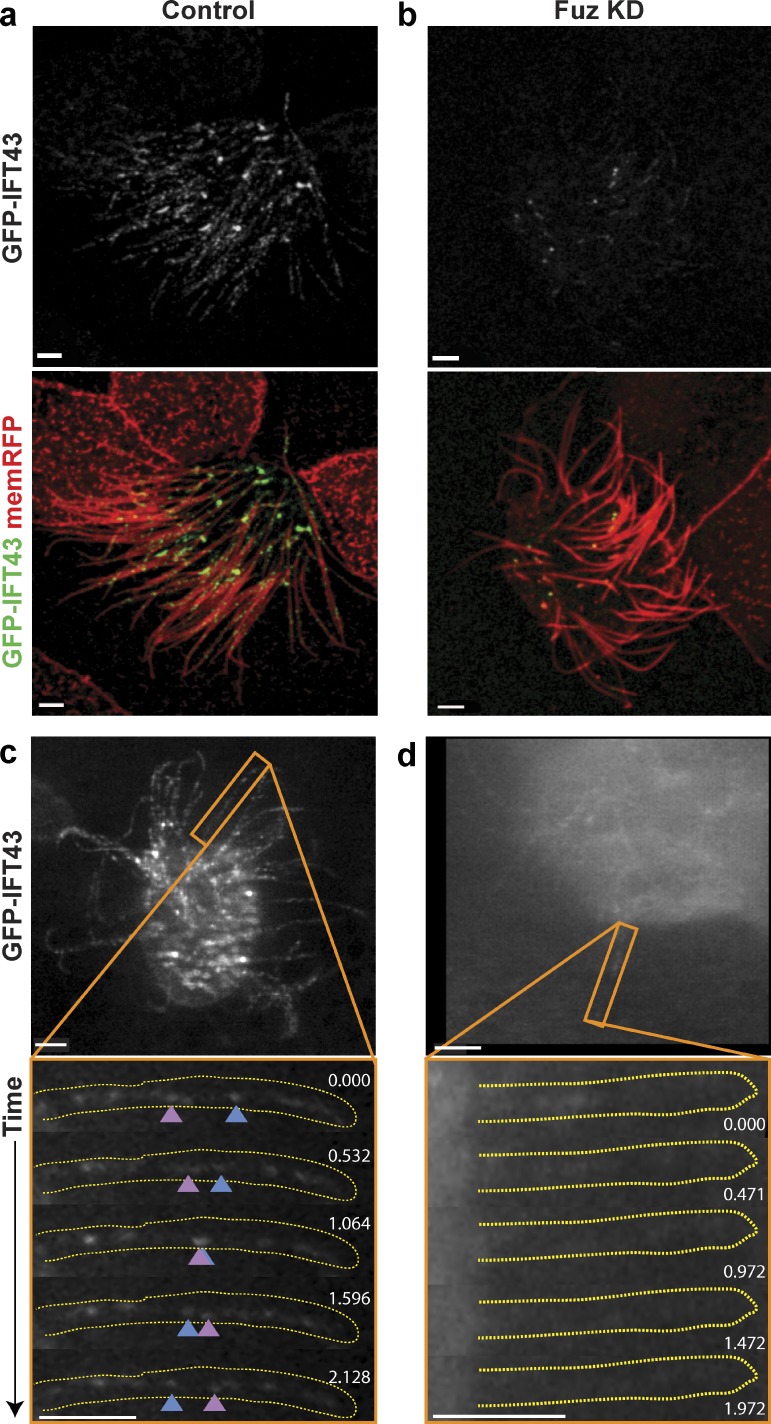
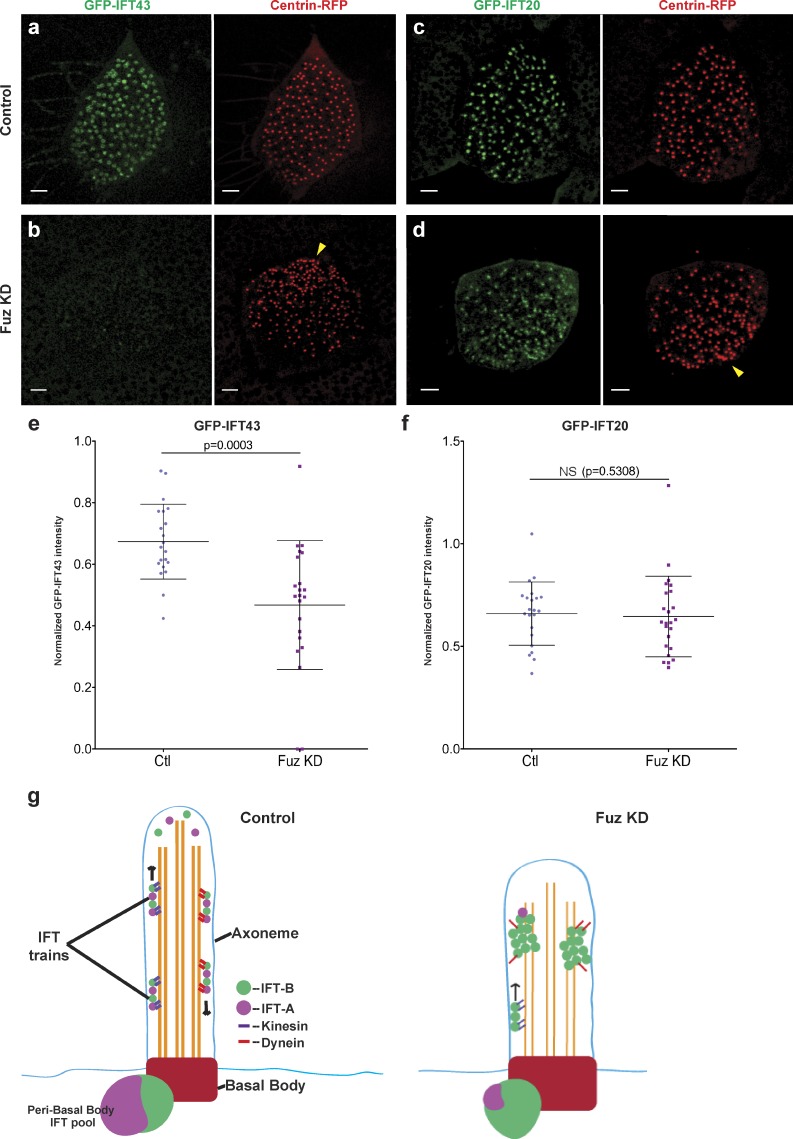
Cilia play key roles in development and homeostasis, and defects in cilia structure or function lead to an array of human diseases. Ciliogenesis is accomplished by the intraflagellar transport (IFT) system, a set of proteins governing bidirectional transport of cargoes within ciliary axonemes. In this paper, we present a novel platform for in vivo analysis of vertebrate IFT dynamics. Using this platform, we show that the planar cell polarity (PCP) effector Fuz was required for normal IFT dynamics in vertebrate cilia, the first evidence directly linking PCP to the core machinery of ciliogenesis. Further, we show that Fuz played a specific role in trafficking of retrograde, but not anterograde, IFT proteins. These data place Fuz in the small group of known IFT effectors outside the core machinery and, additionally, identify Fuz as a novel cytoplasmic effector that differentiates between the retrograde and anterograde IFT complexes.
Figures
Similar articles
-
The Small GTPase Rsg1 is important for the cytoplasmic localization and axonemal dynamics of intraflagellar transport proteins.Cilia. 2013 Oct 7;2:13. doi: 10.1186/2046-2530-2-13. eCollection 2013. Cilia. 2013. PMID: 24192041 Free PMC article.
-
The planar cell polarity effector Fuz is essential for targeted membrane trafficking, ciliogenesis and mouse embryonic development.Nat Cell Biol. 2009 Oct;11(10):1225-32. doi: 10.1038/ncb1966. Epub 2009 Sep 20. Nat Cell Biol. 2009. PMID: 19767740 Free PMC article.
-
The Fuzzy planar cell polarity protein (FUZ), necessary for primary cilium formation, is essential for pituitary development.J Anat. 2024 Feb;244(2):358-367. doi: 10.1111/joa.13961. Epub 2023 Oct 4. J Anat. 2024. PMID: 37794731 Free PMC article.
-
Architecture of the IFT ciliary trafficking machinery and interplay between its components.Crit Rev Biochem Mol Biol. 2020 Apr;55(2):179-196. doi: 10.1080/10409238.2020.1768206. Epub 2020 May 26. Crit Rev Biochem Mol Biol. 2020. PMID: 32456460 Review.
-
Intraflagellar transport (IFT) role in ciliary assembly, resorption and signalling.Curr Top Dev Biol. 2008;85:23-61. doi: 10.1016/S0070-2153(08)00802-8. Curr Top Dev Biol. 2008. PMID: 19147001 Review.
Cited by
-
Total internal reflection fluorescence microscopy of intraflagellar transport in Tetrahymena thermophila.Methods Cell Biol. 2015;127:445-56. doi: 10.1016/bs.mcb.2015.01.001. Epub 2015 Feb 14. Methods Cell Biol. 2015. PMID: 25837403 Free PMC article.
-
Cluap1 is essential for ciliogenesis and photoreceptor maintenance in the vertebrate eye.Invest Ophthalmol Vis Sci. 2014 Jun 26;55(7):4585-92. doi: 10.1167/iovs.14-14888. Invest Ophthalmol Vis Sci. 2014. PMID: 24970261 Free PMC article.
-
Loss of Planar Cell Polarity Effector Fuzzy Causes Renal Hypoplasia by Disrupting Several Signaling Pathways.J Dev Biol. 2021 Dec 23;10(1):1. doi: 10.3390/jdb10010001. J Dev Biol. 2021. PMID: 35076510 Free PMC article.
-
Cilia proteins getting to work - how do they commute from the cytoplasm to the base of cilia?J Cell Sci. 2022 Sep 1;135(17):jcs259444. doi: 10.1242/jcs.259444. Epub 2022 Sep 8. J Cell Sci. 2022. PMID: 36073764 Free PMC article. Review.
-
Coordinated genomic control of ciliogenesis and cell movement by RFX2.Elife. 2014;3:e01439. doi: 10.7554/eLife.01439. Epub 2014 Jan 14. Elife. 2014. PMID: 24424412 Free PMC article.
References
-
- Blacque O.E., Reardon M.J., Li C., McCarthy J., Mahjoub M.R., Ansley S.J., Badano J.L., Mah A.K., Beales P.L., Davidson W.S., et al. 2004. Loss of C. elegans BBS-7 and BBS-8 protein function results in cilia defects and compromised intraflagellar transport. Genes Dev. 18:1630–1642 10.1101/gad.1194004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
