

Transient receptor potential canonical type 3 channels facilitate endothelium-derived hyperpolarization-mediated resistance artery vasodilator activity
- PMID: 22721989
- PMCID: PMC3422079
- DOI: 10.1093/cvr/cvs208
Transient receptor potential canonical type 3 channels facilitate endothelium-derived hyperpolarization-mediated resistance artery vasodilator activity
Abstract
Aims: Microdomain signalling mechanisms underlie key aspects of artery function and the modulation of intracellular calcium, with transient receptor potential (TRP) channels playing an integral role. This study determines the distribution and role of TRP canonical type 3 (C3) channels in the control of endothelium-derived hyperpolarization (EDH)-mediated vasodilator tone in rat mesenteric artery.
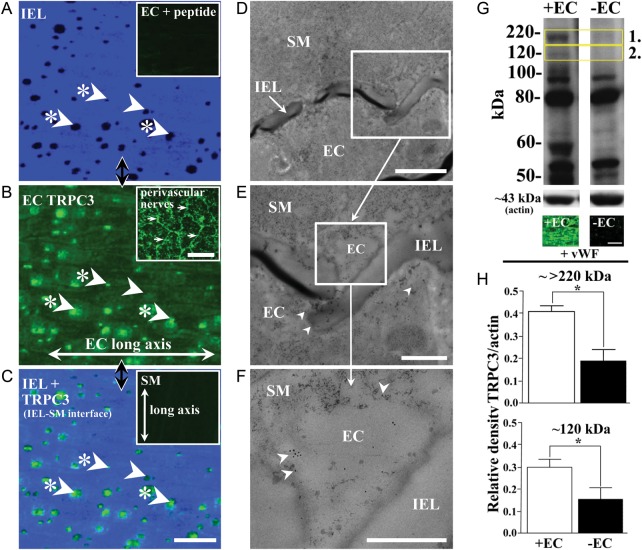
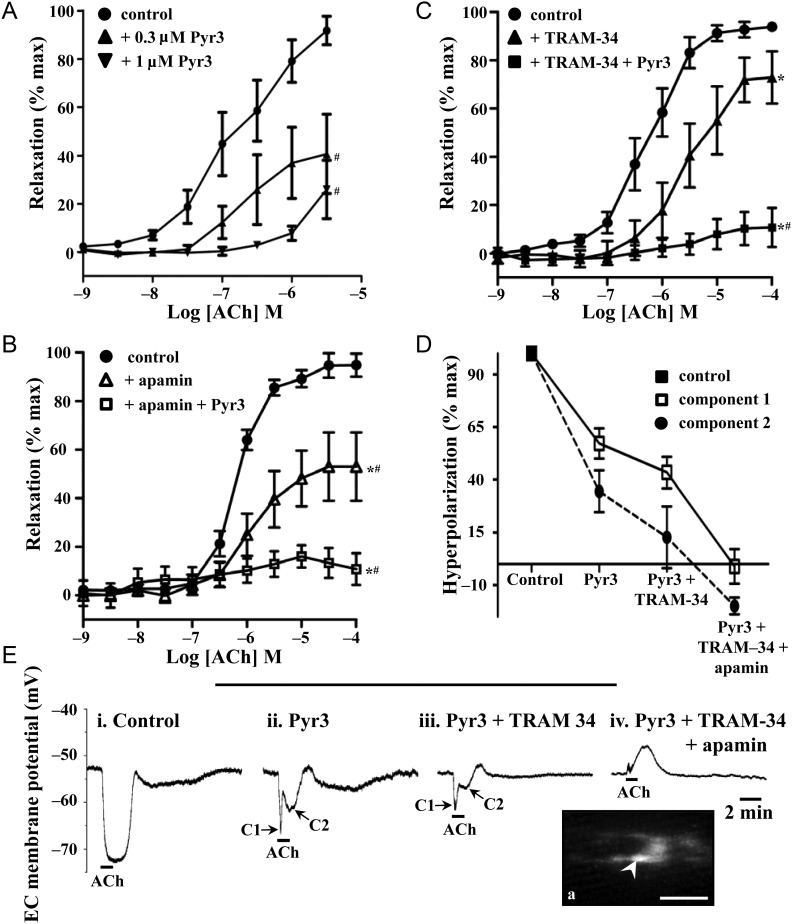
Methods and results: TRPC3 antibody specificity was verified using rat tissue, human embryonic kidney (HEK)-293 cells stably transfected with mouse TRPC3 cDNA, and TRPC3 knock-out (KO) mouse tissue using western blotting and confocal and ultrastructural immunohistochemistry. TRPC3-Pyr3 (ethyl-1-(4-(2,3,3-trichloroacrylamide)phenyl)-5-(trifluoromethyl)-1H-pyrazole-4-carboxylate) specificity was verified using patch clamp of mouse mesenteric artery endothelial and TRPC3-transfected HEK cells, and TRPC3 KO and wild-type mouse aortic endothelial cell calcium imaging and mesenteric artery pressure myography. TRPC3 distribution, expression, and role in EDH-mediated function were examined in rat mesenteric artery using immunohistochemistry and western blotting, and pressure myography and endothelial cell membrane potential recordings. In rat mesenteric artery, TRPC3 was diffusely distributed in the endothelium, with approximately five-fold higher expression at potential myoendothelial microdomain contact sites, and immunoelectron microscopy confirmed TRPC3 at these sites. Western blotting and endothelial damage confirmed primary endothelial TRPC3 expression. In rat mesenteric artery endothelial cells, Pyr3 inhibited hyperpolarization generation, and with individual SK(Ca) (apamin) or IK(Ca) (TRAM-34) block, Pyr3 abolished the residual respective IK(Ca)- and SK(Ca)-dependent EDH-mediated vasodilation.
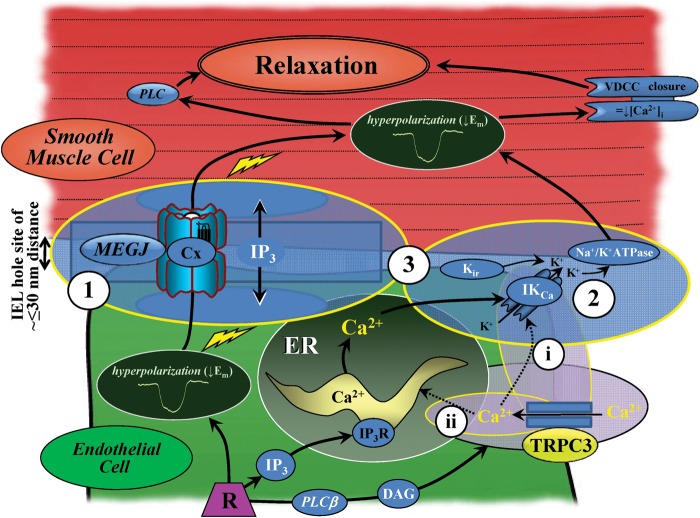
Conclusion: The spatial localization of TRPC3 and associated channels, receptors, and calcium stores are integral for myoendothelial microdomain function. TRPC3 facilitates endothelial SK(Ca) and IK(Ca) activation, as key components of EDH-mediated vasodilator activity and for regulating mesenteric artery tone.
Figures
Comment in
-
Does TRPC3 macrodominate the myoendothelial gap junction microdomain?Cardiovasc Res. 2012 Sep 1;95(4):399-400. doi: 10.1093/cvr/cvs243. Epub 2012 Jul 20. Cardiovasc Res. 2012. PMID: 22822103 No abstract available.
Similar articles
-
Activation of endothelial transient receptor potential C3 channel is required for small conductance calcium-activated potassium channel activation and sustained endothelial hyperpolarization and vasodilation of cerebral artery.J Am Heart Assoc. 2014 Aug 20;3(4):e000913. doi: 10.1161/JAHA.114.000913. J Am Heart Assoc. 2014. PMID: 25142058 Free PMC article.
-
Type 2 diabetes: increased expression and contribution of IKCa channels to vasodilation in small mesenteric arteries of ZDF rats.Am J Physiol Heart Circ Physiol. 2014 Oct 15;307(8):H1093-102. doi: 10.1152/ajpheart.00240.2013. Epub 2014 Aug 15. Am J Physiol Heart Circ Physiol. 2014. PMID: 25128173
-
β₁-Adrenoceptor stimulation suppresses endothelial IK(Ca)-channel hyperpolarization and associated dilatation in resistance arteries.Br J Pharmacol. 2013 Jun;169(4):875-86. doi: 10.1111/bph.12160. Br J Pharmacol. 2013. PMID: 23488860 Free PMC article.
-
Transient receptor potential canonical type 3 channels: Interactions, role and relevance - A vascular focus.Pharmacol Ther. 2017 Jun;174:79-96. doi: 10.1016/j.pharmthera.2017.02.022. Epub 2017 Feb 14. Pharmacol Ther. 2017. PMID: 28223224 Review.
-
Endothelial control of vasodilation: integration of myoendothelial microdomain signalling and modulation by epoxyeicosatrienoic acids.Pflugers Arch. 2014 Mar;466(3):389-405. doi: 10.1007/s00424-013-1303-3. Epub 2013 Jun 8. Pflugers Arch. 2014. PMID: 23748495 Free PMC article. Review.
Cited by
-
Endothelial Ca2+ signaling-dependent vasodilation through transient receptor potential channels.Korean J Physiol Pharmacol. 2020 Jul 1;24(4):287-298. doi: 10.4196/kjpp.2020.24.4.287. Korean J Physiol Pharmacol. 2020. PMID: 32587123 Free PMC article. Review.
-
Transient Receptor Potential Canonical 3 and Nuclear Factor of Activated T Cells C3 Signaling Pathway Critically Regulates Myocardial Fibrosis.Antioxid Redox Signal. 2019 Jun 1;30(16):1851-1879. doi: 10.1089/ars.2018.7545. Epub 2018 Nov 29. Antioxid Redox Signal. 2019. PMID: 30318928 Free PMC article.
-
Calcium signals that determine vascular resistance.Wiley Interdiscip Rev Syst Biol Med. 2019 Sep;11(5):e1448. doi: 10.1002/wsbm.1448. Epub 2019 Mar 18. Wiley Interdiscip Rev Syst Biol Med. 2019. PMID: 30884210 Free PMC article. Review.
-
Endothelium-Dependent Hyperpolarization (EDH) in Hypertension: The Role of Endothelial Ion Channels.Int J Mol Sci. 2018 Jan 21;19(1):315. doi: 10.3390/ijms19010315. Int J Mol Sci. 2018. PMID: 29361737 Free PMC article. Review.
-
Role of TRP channels in the cardiovascular system.Am J Physiol Heart Circ Physiol. 2015 Feb 1;308(3):H157-82. doi: 10.1152/ajpheart.00457.2014. Epub 2014 Nov 21. Am J Physiol Heart Circ Physiol. 2015. PMID: 25416190 Free PMC article. Review.
References
-
- Sandow SL, Tare M. C-type natriuretic peptide: a new endothelium-derived hyperpolarizing factor? Trends Pharmacol Sci. 2007;28:61–67. - PubMed
-
- Kohler R, Hoyer J. The endothelium-derived hyperpolarizing factor: insights from genetic animal models. Kidney Int. 2007;72:145–150. - PubMed
-
- Sandow SL, Tare M, Coleman HA, Hill CE, Parkington HC. Involvement of myoendothelial gap junctions in the actions of EDHF. Circ Res. 2002;90:1108–1113. - PubMed
-
- Mather S, Dora KA, Sandow SL, Winter P, Garland CJ. Rapid endothelial cell-selective loading of connexin 40 antibody blocks EDHF dilation in rat small mesenteric arteries. Circ Res. 2005;97:399–407. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
