

Long noncoding RNAs in C. elegans
- PMID: 22707570
- PMCID: PMC3514682
- DOI: 10.1101/gr.140475.112
Long noncoding RNAs in C. elegans
Abstract
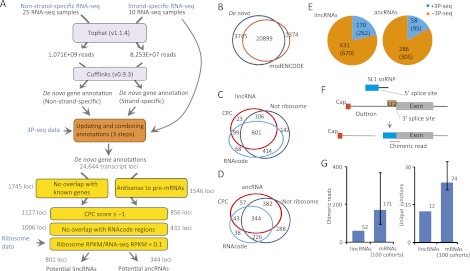
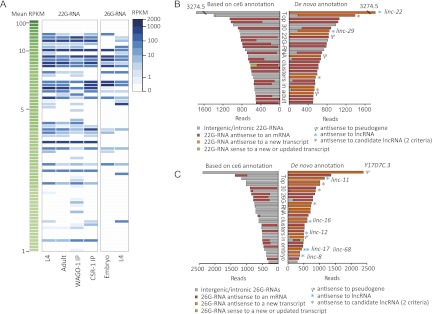
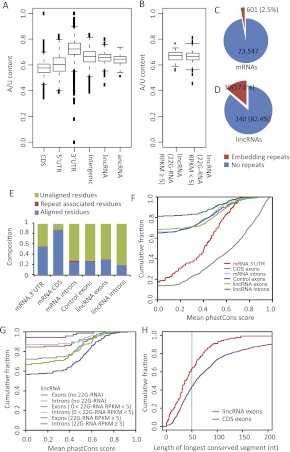
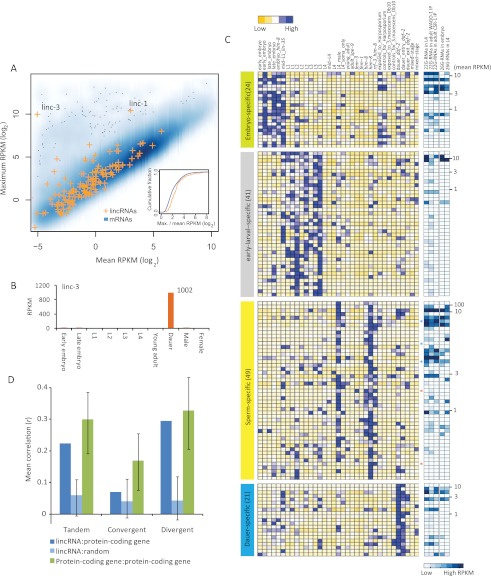
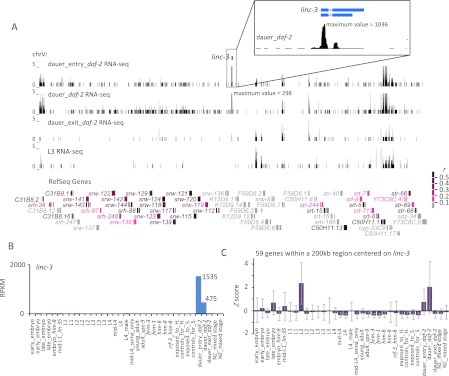
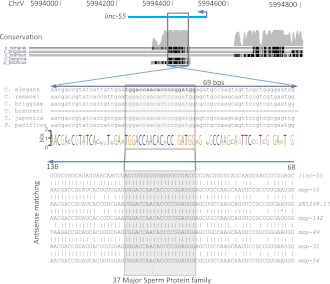
Thousands of long noncoding RNAs (lncRNAs) have been found in vertebrate animals, a few of which have known biological roles. To better understand the genomics and features of lncRNAs in invertebrates, we used available RNA-seq, poly(A)-site, and ribosome-mapping data to identify lncRNAs of Caenorhabditis elegans. We found 170 long intervening ncRNAs (lincRNAs), which had single- or multiexonic structures that did not overlap protein-coding transcripts, and about sixty antisense lncRNAs (ancRNAs), which were complementary to protein-coding transcripts. Compared to protein-coding genes, the lncRNA genes tended to be expressed in a stage-dependent manner. Approximately 25% of the newly identified lincRNAs showed little signal for sequence conservation and mapped antisense to clusters of endogenous siRNAs, as would be expected if they serve as templates and targets for these siRNAs. The other 75% tended to be more conserved and included lincRNAs with intriguing expression and sequence features associating them with processes such as dauer formation, male identity, sperm formation, and interaction with sperm-specific mRNAs. Our study provides a glimpse into the lncRNA content of a nonvertebrate animal and a resource for future studies of lncRNA function.
Figures
Similar articles
-
Systematic identification of long intergenic non-coding RNAs expressed in bovine oocytes.Reprod Biol Endocrinol. 2020 Feb 21;18(1):13. doi: 10.1186/s12958-020-00573-4. Reprod Biol Endocrinol. 2020. PMID: 32085734 Free PMC article.
-
Bioinformatics Approaches to Studying Plant Long Noncoding RNAs (lncRNAs): Identification and Functional Interpretation of lncRNAs from RNA-Seq Data Sets.Methods Mol Biol. 2019;1933:197-205. doi: 10.1007/978-1-4939-9045-0_11. Methods Mol Biol. 2019. PMID: 30945186
-
Splicing conservation signals in plant long noncoding RNAs.RNA. 2020 Jul;26(7):784-793. doi: 10.1261/rna.074393.119. Epub 2020 Apr 2. RNA. 2020. PMID: 32241834 Free PMC article.
-
Roles of long noncoding RNAs in brain development, functional diversification and neurodegenerative diseases.Brain Res Bull. 2013 Aug;97:69-80. doi: 10.1016/j.brainresbull.2013.06.001. Epub 2013 Jun 10. Brain Res Bull. 2013. PMID: 23756188 Review.
-
Long noncoding RNAs: Novel insights into hepatocelluar carcinoma.Cancer Lett. 2014 Mar 1;344(1):20-27. doi: 10.1016/j.canlet.2013.10.021. Epub 2013 Oct 30. Cancer Lett. 2014. PMID: 24183851 Review.
Cited by
-
The physiological function of long-noncoding RNAs.Noncoding RNA Res. 2020 Dec;5(4):178-184. doi: 10.1016/j.ncrna.2020.09.003. Epub 2020 Sep 17. Noncoding RNA Res. 2020. PMID: 32959025 Free PMC article. Review.
-
Genome-wide analysis of transcription-coupled repair reveals novel transcription events in Caenorhabditis elegans.PLoS Genet. 2024 Jul 19;20(7):e1011365. doi: 10.1371/journal.pgen.1011365. eCollection 2024 Jul. PLoS Genet. 2024. PMID: 39028758 Free PMC article.
-
Divergent transcription is associated with promoters of transcriptional regulators.BMC Genomics. 2013 Dec 23;14:914. doi: 10.1186/1471-2164-14-914. BMC Genomics. 2013. PMID: 24365181 Free PMC article.
-
Genome and transcriptome of the porcine whipworm Trichuris suis.Nat Genet. 2014 Jul;46(7):701-6. doi: 10.1038/ng.3012. Epub 2014 Jun 15. Nat Genet. 2014. PMID: 24929829 Free PMC article.
-
The time-resolved transcriptome of C. elegans.Genome Res. 2016 Oct;26(10):1441-1450. doi: 10.1101/gr.202663.115. Epub 2016 Aug 16. Genome Res. 2016. PMID: 27531719 Free PMC article.
References
-
- Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D 2003. MicroRNAs and other tiny endogenous RNAs in C. elegans. Curr Biol 13: 807–818 - PubMed
-
- Blumenthal T, Gleason KS 2003. Caenorhabditis elegans operons: Form and function. Nat Rev Genet 4: 112–120 - PubMed
-
- Blumenthal T, Steward K 1997. RNA processing and gene structure. In C. elegans II (ed. DL Riddle et al.), pp. 117–145. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases