

p53 dynamics control cell fate
- PMID: 22700930
- PMCID: PMC4162876
- DOI: 10.1126/science.1218351
p53 dynamics control cell fate
Abstract
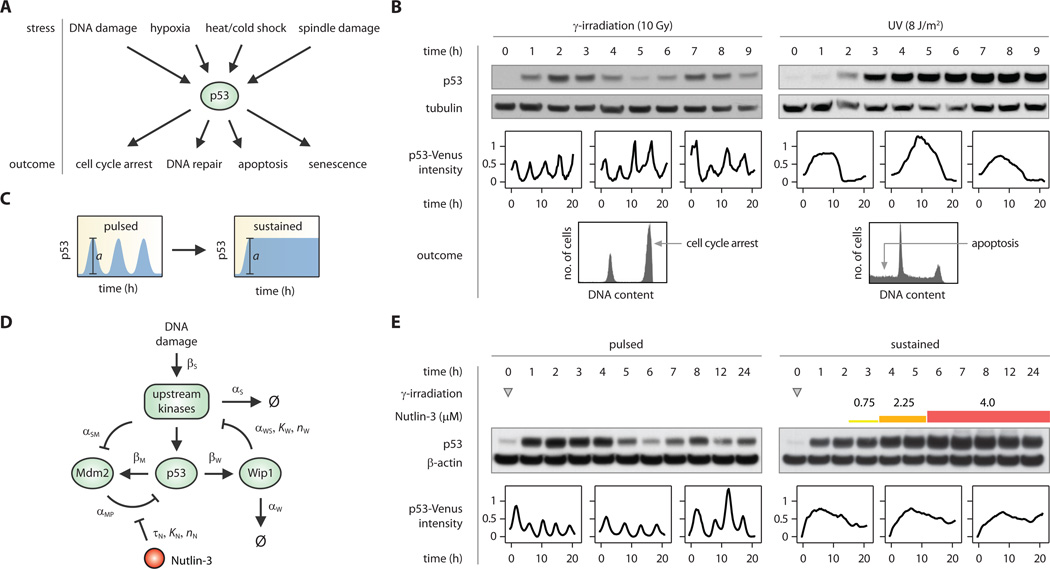
Cells transmit information through molecular signals that often show complex dynamical patterns. The dynamic behavior of the tumor suppressor p53 varies depending on the stimulus; in response to double-strand DNA breaks, it shows a series of repeated pulses. Using a computational model, we identified a sequence of precisely timed drug additions that alter p53 pulses to instead produce a sustained p53 response. This leads to the expression of a different set of downstream genes and also alters cell fate: Cells that experience p53 pulses recover from DNA damage, whereas cells exposed to sustained p53 signaling frequently undergo senescence. Our results show that protein dynamics can be an important part of a signal, directly influencing cellular fate decisions.
Figures



Similar articles
-
p53 and p21(Waf1) are recruited to distinct PML-containing nuclear foci in irradiated and Nutlin-3a-treated U2OS cells.J Cell Biochem. 2010 Dec 1;111(5):1280-90. doi: 10.1002/jcb.22852. J Cell Biochem. 2010. PMID: 20803550 Free PMC article.
-
Cell fate decision mediated by p53 pulses.Proc Natl Acad Sci U S A. 2009 Jul 28;106(30):12245-50. doi: 10.1073/pnas.0813088106. Epub 2009 Jul 15. Proc Natl Acad Sci U S A. 2009. PMID: 19617533 Free PMC article.
-
c-ABL tyrosine kinase modulates p53-dependent p21 induction and ensuing cell fate decision in response to DNA damage.Cell Signal. 2014 Feb;26(2):444-52. doi: 10.1016/j.cellsig.2013.10.005. Epub 2013 Oct 28. Cell Signal. 2014. PMID: 24177958
-
Significance of Wild-Type p53 Signaling in Suppressing Apoptosis in Response to Chemical Genotoxic Agents: Impact on Chemotherapy Outcome.Int J Mol Sci. 2017 Apr 28;18(5):928. doi: 10.3390/ijms18050928. Int J Mol Sci. 2017. PMID: 28452953 Free PMC article. Review.
-
Progress and challenges in understanding the regulation and function of p53 dynamics.Biochem Soc Trans. 2021 Nov 1;49(5):2123-2131. doi: 10.1042/BST20210148. Biochem Soc Trans. 2021. PMID: 34495325 Free PMC article. Review.
Cited by
-
Beyond generalized hair cells: molecular cues for hair cell types.Hear Res. 2013 Mar;297:30-41. doi: 10.1016/j.heares.2012.11.008. Epub 2012 Nov 27. Hear Res. 2013. PMID: 23201032 Free PMC article. Review.
-
A Cre-dependent reporter mouse for quantitative real-time imaging of protein kinase A activity dynamics.Sci Rep. 2024 Feb 6;14(1):3054. doi: 10.1038/s41598-024-53313-8. Sci Rep. 2024. PMID: 38321128 Free PMC article.
-
Coordination of MAPK and p53 dynamics in the cellular responses to DNA damage and oxidative stress.Mol Syst Biol. 2022 Dec;18(12):e11401. doi: 10.15252/msb.202211401. Mol Syst Biol. 2022. PMID: 36472304 Free PMC article.
-
Tumor suppressor p53 plays a key role in induction of both tristetraprolin and let-7 in human cancer cells.Nucleic Acids Res. 2013 Jun;41(11):5614-25. doi: 10.1093/nar/gkt222. Epub 2013 Apr 16. Nucleic Acids Res. 2013. PMID: 23595149 Free PMC article.
-
Emerging role and therapeutic implications of p53 in intervertebral disc degeneration.Cell Death Discov. 2023 Dec 1;9(1):433. doi: 10.1038/s41420-023-01730-5. Cell Death Discov. 2023. PMID: 38040675 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
