

Influence of herpes simplex virus 1 latency-associated transcripts on the establishment and maintenance of latency in the ROSA26R reporter mouse model
- PMID: 22696655
- PMCID: PMC3421754
- DOI: 10.1128/JVI.00652-12
Influence of herpes simplex virus 1 latency-associated transcripts on the establishment and maintenance of latency in the ROSA26R reporter mouse model
Abstract
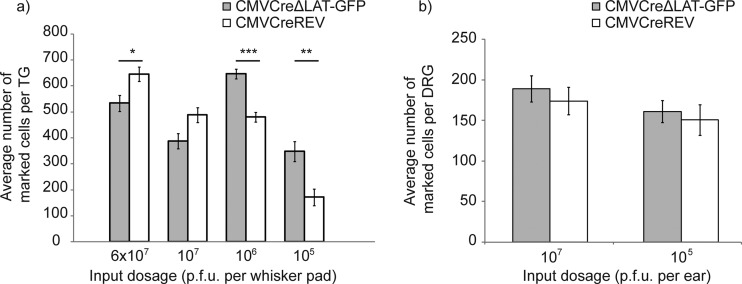
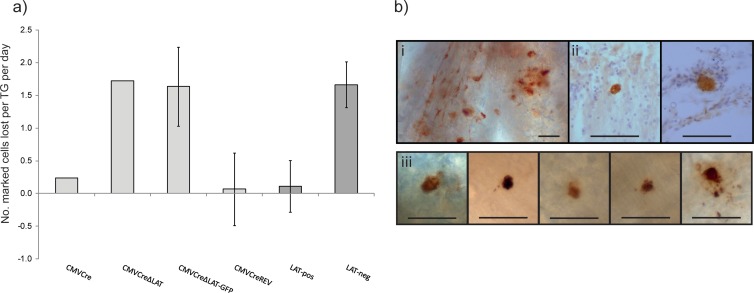
Herpes simplex virus 1 (HSV-1) can establish life-long latent infection in sensory neurons, from which periodic reactivation can occur. During latency, viral gene expression is largely restricted to the latency-associated transcripts (LATs). While not essential for any phase of latency, to date the LATs have been shown to increase the efficiency of both establishment and reactivation of latency in small-animal models. We sought to investigate the role of LAT expression in the frequency of latency establishment within the ROSA26R reporter mouse model utilizing Cre recombinase-encoding recombinant viruses harboring deletions of the core LAT promoter (LAP) region. HSV-1 LAT expression was observed to influence the number of latently infected neurons in trigeminal but not dorsal root ganglia. Furthermore, the relative frequencies of latency establishment of LAT-positive and LAT-negative viruses are influenced by the inoculum dose following infection of the mouse whisker pads. Finally, analysis of the infected cell population at two latent time points revealed a relative loss of latently infected cells in the absence of LAT expression. We conclude that the HSV-1 LATs facilitate the long-term stability of the latent cell population within the infected host and that interpretation of LAT establishment phenotypes is influenced by infection methodology.
Figures
Similar articles
-
Herpes simplex virus type 2 (HSV-2) establishes latent infection in a different population of ganglionic neurons than HSV-1: role of latency-associated transcripts.J Virol. 2007 Feb;81(4):1872-8. doi: 10.1128/JVI.02110-06. Epub 2006 Dec 6. J Virol. 2007. PMID: 17151134 Free PMC article.
-
Herpes Simplex Virus Type 1 Preferentially Enhances Neuro-Inflammation and Senescence in Brainstem of Female Mice.J Virol. 2022 Sep 14;96(17):e0108122. doi: 10.1128/jvi.01081-22. Epub 2022 Aug 17. J Virol. 2022. PMID: 35975996 Free PMC article.
-
In Vivo Knockdown of the Herpes Simplex Virus 1 Latency-Associated Transcript Reduces Reactivation from Latency.J Virol. 2018 Jul 31;92(16):e00812-18. doi: 10.1128/JVI.00812-18. Print 2018 Aug 15. J Virol. 2018. PMID: 29875240 Free PMC article.
-
A comparison of herpes simplex virus type 1 and varicella-zoster virus latency and reactivation.J Gen Virol. 2015 Jul;96(Pt 7):1581-602. doi: 10.1099/vir.0.000128. Epub 2015 Mar 20. J Gen Virol. 2015. PMID: 25794504 Free PMC article. Review.
-
Molecular circuitry regulating herpes simplex virus type 1 latency in neurons.J Neurovirol. 2000 Feb;6(1):6-24. doi: 10.3109/13550280009006378. J Neurovirol. 2000. PMID: 10786993 Review.
Cited by
-
The HSV-1 Latency-Associated Transcript Functions to Repress Latent Phase Lytic Gene Expression and Suppress Virus Reactivation from Latently Infected Neurons.PLoS Pathog. 2016 Apr 7;12(4):e1005539. doi: 10.1371/journal.ppat.1005539. eCollection 2016 Apr. PLoS Pathog. 2016. PMID: 27055281 Free PMC article.
-
Expression of the herpes simplex virus type 1 latency-associated transcripts does not influence latency establishment of virus mutants deficient for neuronal replication.J Gen Virol. 2013 Nov;94(Pt 11):2489-2494. doi: 10.1099/vir.0.056176-0. Epub 2013 Aug 1. J Gen Virol. 2013. PMID: 23907392 Free PMC article.
-
An Immortalized Human Dorsal Root Ganglion Cell Line Provides a Novel Context To Study Herpes Simplex Virus 1 Latency and Reactivation.J Virol. 2017 May 26;91(12):e00080-17. doi: 10.1128/JVI.00080-17. Print 2017 Jun 15. J Virol. 2017. PMID: 28404842 Free PMC article.
-
Detection of the genome and transcripts of a persistent DNA virus in neuronal tissues by fluorescent in situ hybridization combined with immunostaining.J Vis Exp. 2014 Jan 23;(83):e51091. doi: 10.3791/51091. J Vis Exp. 2014. PMID: 24514006 Free PMC article.
-
Sex differences underlying orofacial varicella zoster associated pain in rats.BMC Neurol. 2017 May 17;17(1):95. doi: 10.1186/s12883-017-0882-6. BMC Neurol. 2017. PMID: 28514943 Free PMC article.
References
-
- Arthur JL, Everett R, Brierley I, Efstathiou S. 1998. Disruption of the 5′ and 3′ splice sites flanking the major latency-associated transcripts of herpes simplex virus type 1: evidence for alternate splicing in lytic and latent infections. J. Gen. Virol. 79(Pt 1):107–116 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
