

Caspase-11 promotes the fusion of phagosomes harboring pathogenic bacteria with lysosomes by modulating actin polymerization
- PMID: 22658523
- PMCID: PMC3408798
- DOI: 10.1016/j.immuni.2012.05.001
Caspase-11 promotes the fusion of phagosomes harboring pathogenic bacteria with lysosomes by modulating actin polymerization
Abstract
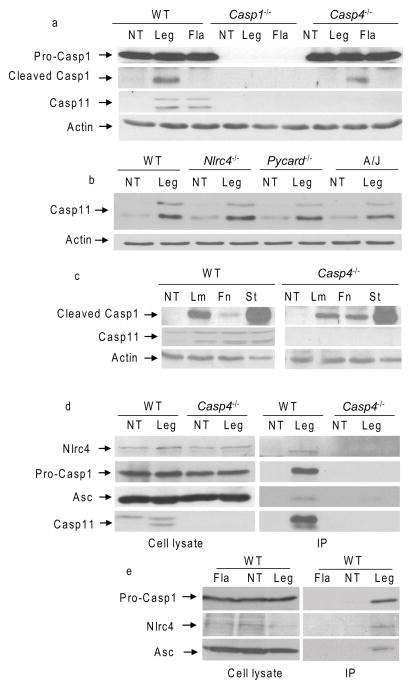
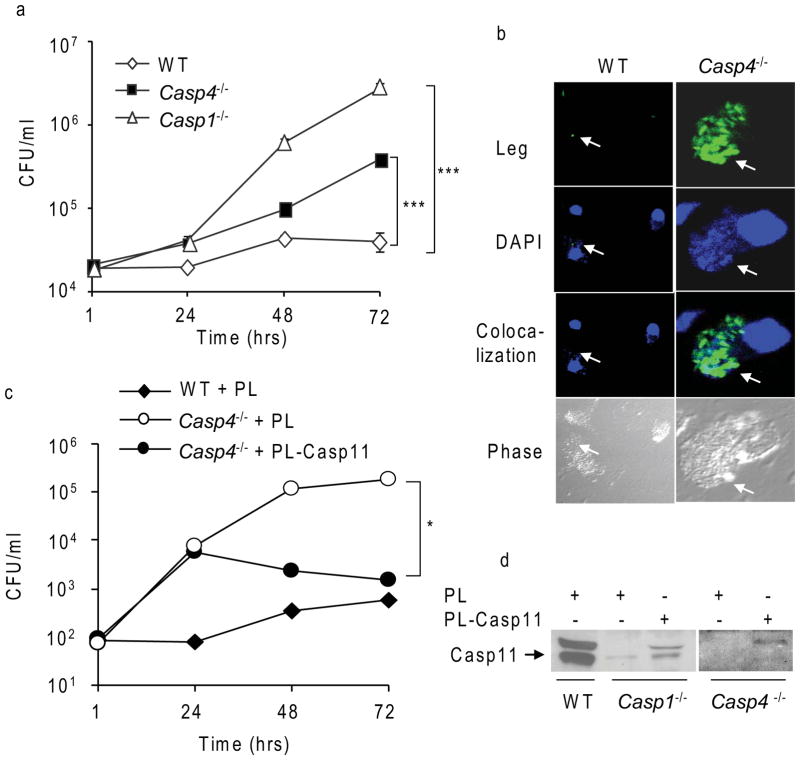
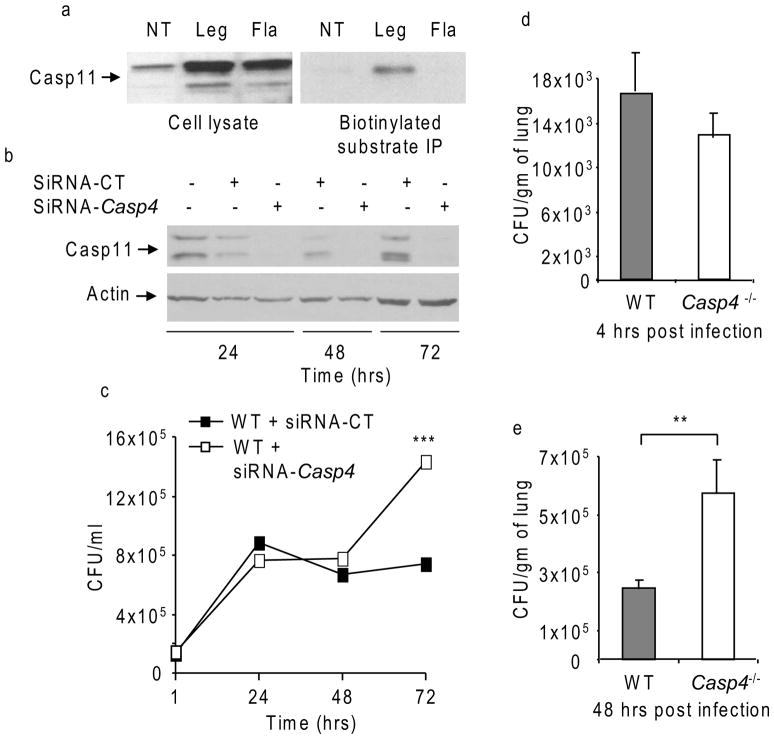
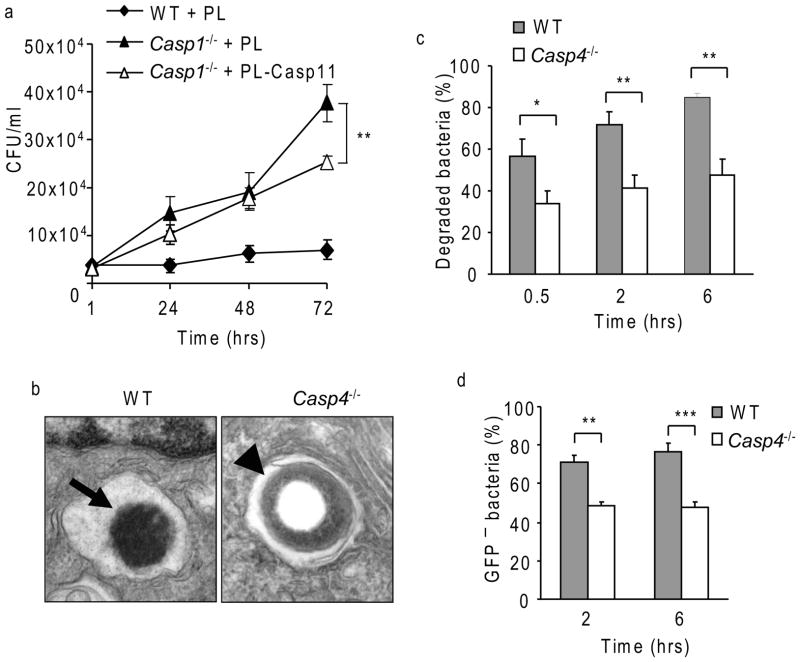
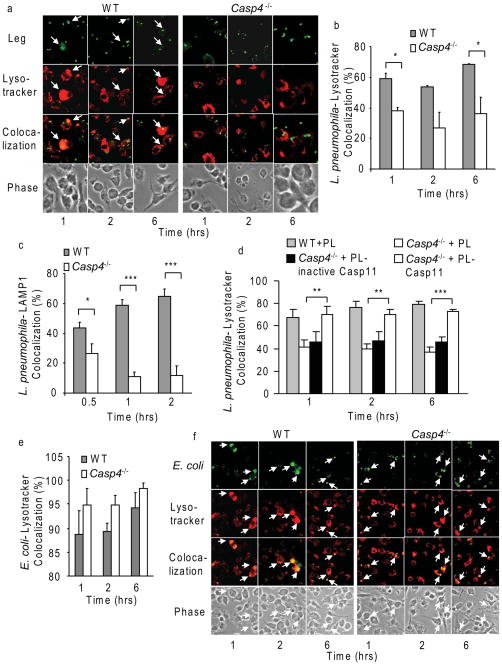
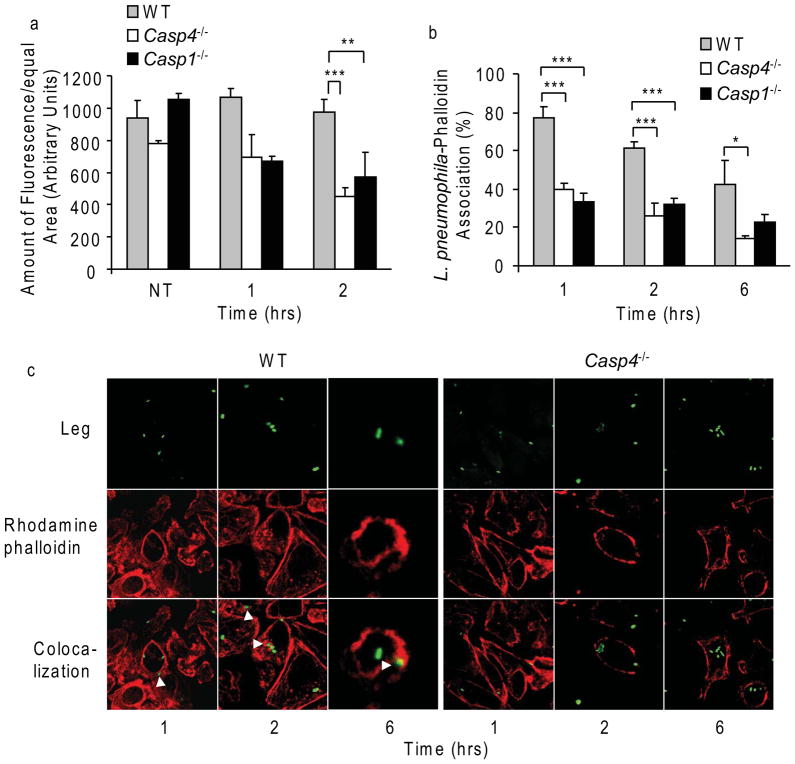
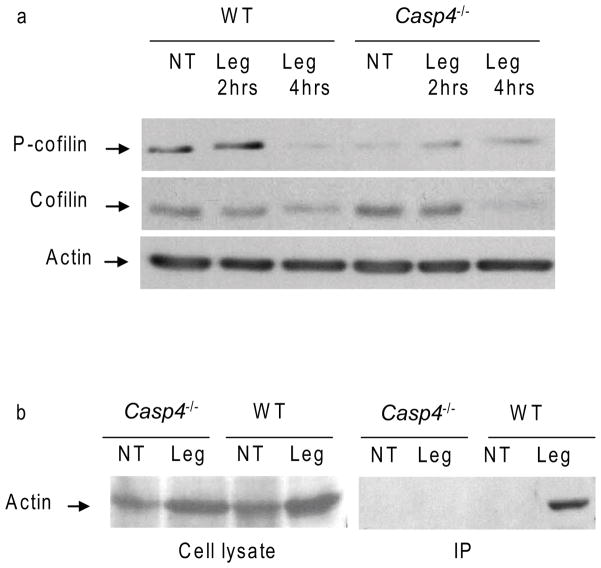
Inflammasomes are multiprotein complexes that include members of the NLR (nucleotide-binding domain leucine-rich repeat containing) family and caspase-1. Once bacterial molecules are sensed within the macrophage, the inflammasome is assembled, mediating the activation of caspase-1. Caspase-11 mediates caspase-1 activation in response to lipopolysaccharide and bacterial toxins, and yet its role during bacterial infection is unknown. Here, we demonstrated that caspase-11 was dispensable for caspase-1 activation in response to Legionella, Salmonella, Francisella, and Listeria. We also determined that active mouse caspase-11 was required for restriction of L. pneumophila infection. Similarly, human caspase-4 and caspase-5, homologs of mouse caspase-11, cooperated to restrict L. pneumophila infection in human macrophages. Caspase-11 promoted the fusion of the L. pneumophila vacuole with lysosomes by modulating actin polymerization through cofilin. However, caspase-11 was dispensable for the fusion of lysosomes with phagosomes containing nonpathogenic bacteria, uncovering a fundamental difference in the trafficking of phagosomes according to their cargo.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
CASP4/caspase-11 promotes autophagosome formation in response to bacterial infection.Autophagy. 2018;14(11):1928-1942. doi: 10.1080/15548627.2018.1491494. Epub 2018 Aug 31. Autophagy. 2018. PMID: 30165781 Free PMC article.
-
The Sphingosine-1-Phosphate Lyase (LegS2) Contributes to the Restriction of Legionella pneumophila in Murine Macrophages.PLoS One. 2016 Jan 7;11(1):e0146410. doi: 10.1371/journal.pone.0146410. eCollection 2016. PLoS One. 2016. PMID: 26741365 Free PMC article.
-
Caspase-11 and caspase-1 differentially modulate actin polymerization via RhoA and Slingshot proteins to promote bacterial clearance.Sci Rep. 2015 Dec 21;5:18479. doi: 10.1038/srep18479. Sci Rep. 2015. PMID: 26686473 Free PMC article.
-
Modulation of caspases and their non-apoptotic functions by Legionella pneumophila.Cell Microbiol. 2010 Feb;12(2):140-7. doi: 10.1111/j.1462-5822.2009.01401.x. Epub 2009 Oct 27. Cell Microbiol. 2010. PMID: 19863553 Review.
-
Caspase-11: the driving factor for noncanonical inflammasomes.Eur J Immunol. 2013 Sep;43(9):2240-5. doi: 10.1002/eji.201343800. Eur J Immunol. 2013. PMID: 24037676 Review.
Cited by
-
Regulation of inflammasome activation.Immunol Rev. 2015 May;265(1):6-21. doi: 10.1111/imr.12296. Immunol Rev. 2015. PMID: 25879280 Free PMC article. Review.
-
Cutting Edge: Caspase-11 Limits the Response of CD8+ T Cells to Low-Abundance and Low-Affinity Antigens.J Immunol. 2015 Jul 1;195(1):41-5. doi: 10.4049/jimmunol.1500812. Epub 2015 May 15. J Immunol. 2015. PMID: 25980012 Free PMC article.
-
Caspase-11 counteracts mitochondrial ROS-mediated clearance of Staphylococcus aureus in macrophages.EMBO Rep. 2019 Dec 5;20(12):e48109. doi: 10.15252/embr.201948109. Epub 2019 Oct 21. EMBO Rep. 2019. PMID: 31637841 Free PMC article.
-
Caspase-4 Mediates Restriction of Burkholderia pseudomallei in Human Alveolar Epithelial Cells.Infect Immun. 2020 Feb 20;88(3):e00868-19. doi: 10.1128/IAI.00868-19. Print 2020 Feb 20. Infect Immun. 2020. PMID: 31818963 Free PMC article.
-
Multiple roles of caspase-8 in cell death, inflammation, and innate immunity.J Leukoc Biol. 2021 Jan;109(1):121-141. doi: 10.1002/JLB.3MR0420-305R. Epub 2020 Jun 12. J Leukoc Biol. 2021. PMID: 32531842 Free PMC article. Review.
References
-
- Abdulrahman BA, Abu Khweek A, Akhter A, Caution K, Kotrange S, Abdelaziz DHA, Newland C, Rosales-Reyes R, Kopp B, McCoy K, et al. Autophagy stimulation by Rapamycin suppresses lung inflammation and infection by Burkholderia cenocepacia in a model of cystic fibrosis Autophagy. 2011. p. 7. - PMC - PubMed
-
- Amer A, Franchi L, Kanneganti TD, Body-Malapel M, Ozoren N, Brady G, Meshinchi S, Jagirdar R, Gewirtz A, Akira S, et al. Regulation of Legionella phagosome maturation and infection through flagellin and host Ipaf. J Biol Chem. 2006;281:35217–35223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
