

Efficient human cytomegalovirus reactivation is maturation dependent in the Langerhans dendritic cell lineage and can be studied using a CD14+ experimental latency model
- PMID: 22647696
- PMCID: PMC3421708
- DOI: 10.1128/JVI.00598-12
Efficient human cytomegalovirus reactivation is maturation dependent in the Langerhans dendritic cell lineage and can be studied using a CD14+ experimental latency model
Abstract
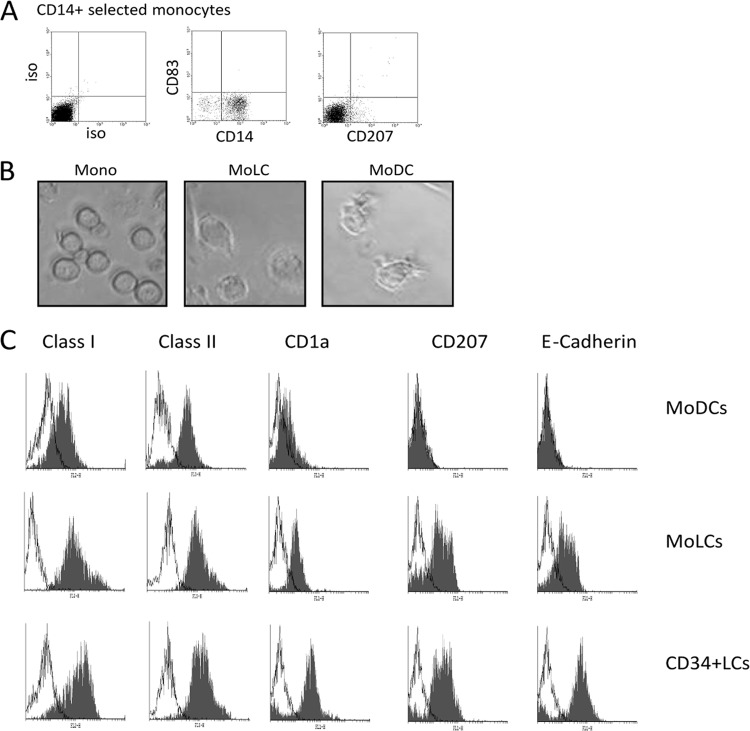
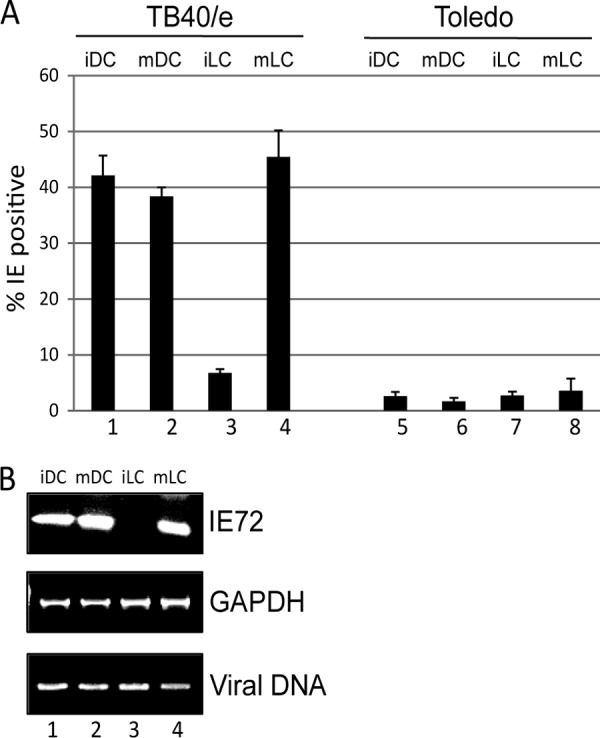
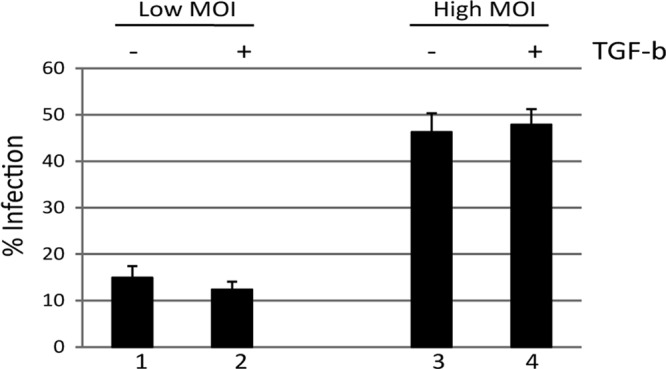
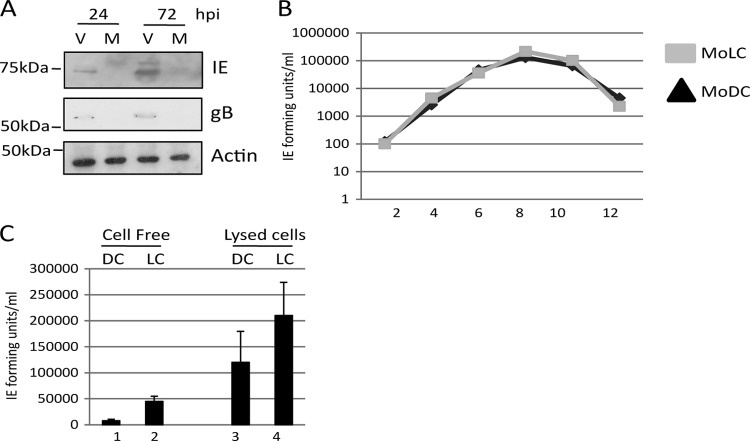
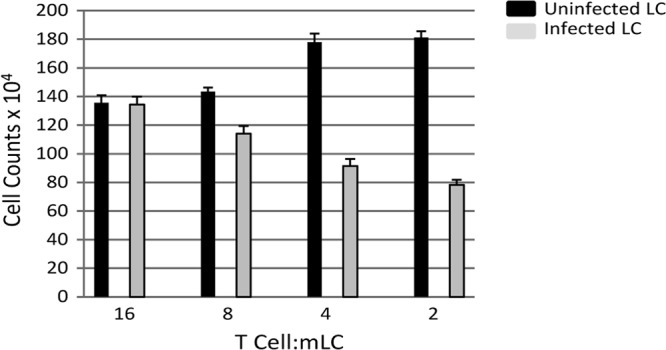
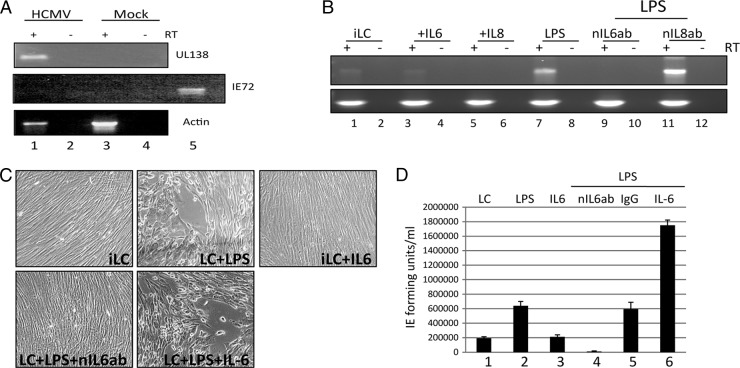
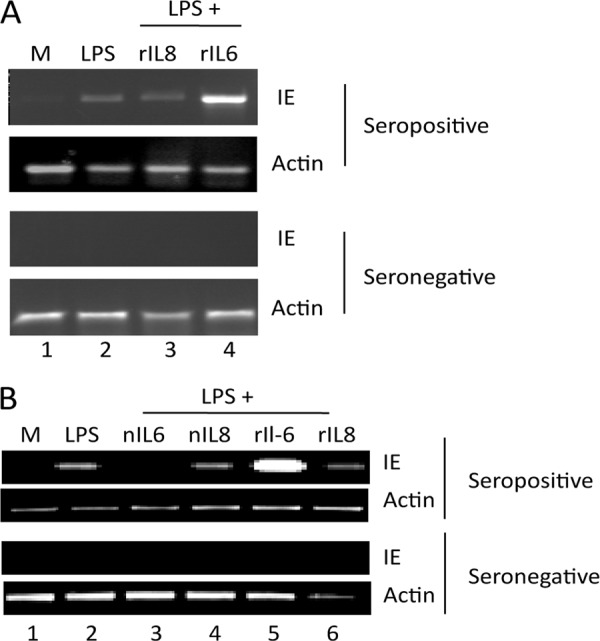
Studies from a number of laboratories have shown that the myeloid lineage is prominent in human cytomegalovirus (HCMV) latency, reactivation, dissemination, and pathogenesis. Existing as a latent infection in CD34(+) progenitors and circulating CD14(+) monocytes, reactivation is observed upon differentiation to mature macrophage or dendritic cell (DC) phenotypes. Langerhans' cells (LCs) are a subset of periphery resident DCs that represent a DC population likely to encounter HCMV early during primary infection. Furthermore, we have previously shown that CD34(+) derived LCs are a site of HCMV reactivation ex vivo. Accordingly, we have utilized healthy-donor CD34(+) cells to study latency and reactivation of HCMV in LCs. However, the increasing difficulty acquiring healthy-donor CD34(+) cells--particularly from seropositive donors due to the screening regimens used--led us to investigate the use of CD14(+) monocytes to generate LCs. We show here that CD14(+) monocytes cultured with transforming growth factor β generate Langerin-positive DCs (MoLCs). Consistent with observations using CD34(+) derived LCs, only mature MoLCs were permissive for HCMV infection. The lytic infection of mature MoLCs is productive and results in a marked inhibition in the capacity of these cells to promote T cell proliferation. Pertinently, differentiation of experimentally latent monocytes to the MoLC phenotype promotes reactivation in a maturation and interleukin-6 (IL-6)-dependent manner. Intriguingly, however, IL-6-mediated effects were restricted to mature LCs, in contrast to observations with classical CD14(+) derived DCs. Consequently, elucidation of the molecular basis behind the differential response of the two DC subsets should further our understanding of the fundamental mechanisms important for reactivation.
Figures

Similar articles
-
Latent Cytomegalovirus-Driven Recruitment of Activated CD4+ T Cells Promotes Virus Reactivation.Front Immunol. 2021 Apr 12;12:657945. doi: 10.3389/fimmu.2021.657945. eCollection 2021. Front Immunol. 2021. PMID: 33912186 Free PMC article.
-
Circulating dendritic cells isolated from healthy seropositive donors are sites of human cytomegalovirus reactivation in vivo.J Virol. 2013 Oct;87(19):10660-7. doi: 10.1128/JVI.01539-13. Epub 2013 Jul 24. J Virol. 2013. PMID: 23885077 Free PMC article.
-
Inhibition of inflammatory interleukin-6 activity via extracellular signal-regulated kinase-mitogen-activated protein kinase signaling antagonizes human cytomegalovirus reactivation from dendritic cells.J Virol. 2011 Dec;85(23):12750-8. doi: 10.1128/JVI.05878-11. Epub 2011 Sep 21. J Virol. 2011. PMID: 21937636 Free PMC article.
-
Human cytomegalovirus: Latency and reactivation in the myeloid lineage.J Clin Virol. 2008 Mar;41(3):180-5. doi: 10.1016/j.jcv.2007.11.014. J Clin Virol. 2008. PMID: 18164651 Review.
-
[Latency and reactivation of HCMV].Nihon Rinsho. 2006 Mar;64 Suppl 3:435-9. Nihon Rinsho. 2006. PMID: 16615510 Review. Japanese. No abstract available.
Cited by
-
Mitogen and stress activated kinases act co-operatively with CREB during the induction of human cytomegalovirus immediate-early gene expression from latency.PLoS Pathog. 2014 Jun 12;10(6):e1004195. doi: 10.1371/journal.ppat.1004195. eCollection 2014 Jun. PLoS Pathog. 2014. PMID: 24945302 Free PMC article.
-
Functional annotation of human cytomegalovirus gene products: an update.Front Microbiol. 2014 May 19;5:218. doi: 10.3389/fmicb.2014.00218. eCollection 2014. Front Microbiol. 2014. PMID: 24904534 Free PMC article. Review.
-
The Expression of Human Cytomegalovirus MicroRNA MiR-UL148D during Latent Infection in Primary Myeloid Cells Inhibits Activin A-triggered Secretion of IL-6.Sci Rep. 2016 Aug 5;6:31205. doi: 10.1038/srep31205. Sci Rep. 2016. PMID: 27491954 Free PMC article.
-
Tumor Necrosis Factor Alpha Induces Reactivation of Human Cytomegalovirus Independently of Myeloid Cell Differentiation following Posttranscriptional Establishment of Latency.mBio. 2018 Sep 11;9(5):e01560-18. doi: 10.1128/mBio.01560-18. mBio. 2018. PMID: 30206173 Free PMC article.
-
Human cytomegalovirus infection of langerhans-type dendritic cells does not require the presence of the gH/gL/UL128-131A complex and is blocked after nuclear deposition of viral genomes in immature cells.J Virol. 2014 Jan;88(1):403-16. doi: 10.1128/JVI.03062-13. Epub 2013 Oct 23. J Virol. 2014. PMID: 24155395 Free PMC article.
References
-
- Adler SP, Hur JK, Wang JB, Vetrovec GW. 1998. Prior infection with cytomegalovirus is not a major risk factor for angiographically demonstrated coronary artery atherosclerosis. J. Infect. Dis. 177:209–212 - PubMed
-
- Allan RS, et al. 2003. Epidermal viral immunity induced by CD8α+ dendritic cells but not by Langerhans cells. Science 301:1925–1928 - PubMed
-
- Bechetoille N, Andre V, Valladeau J, Perrier E, Dezutter-Dambuyant C. 2006. Mixed Langerhans cell and interstitial/dermal dendritic cell subsets emanating from monocytes in Th2-mediated inflammatory conditions respond differently to proinflammatory stimuli. J. Leukoc. Biol. 80:45–58 - PubMed
-
- Bosnjak L, et al. 2005. Herpes simplex virus infection of human dendritic cells induces apoptosis and allows cross-presentation via uninfected dendritic cells. J. Immunol. 174:2220–2227 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
