

Non-Canonical Processing of Arabidopsis pri-miR319a/b/c Generates Additional microRNAs to Target One RAP2.12 mRNA Isoform
- PMID: 22639648
- PMCID: PMC3355612
- DOI: 10.3389/fpls.2012.00046
Non-Canonical Processing of Arabidopsis pri-miR319a/b/c Generates Additional microRNAs to Target One RAP2.12 mRNA Isoform
Abstract
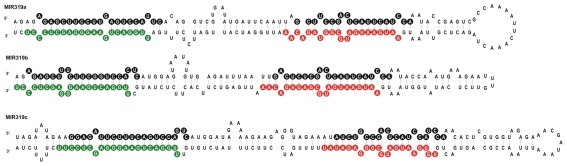
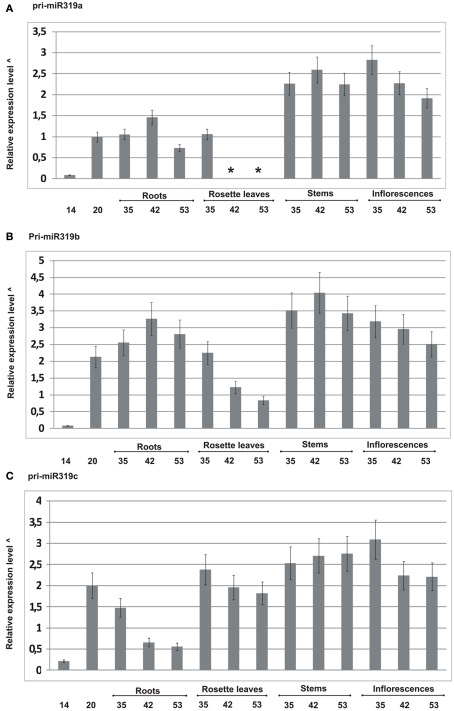
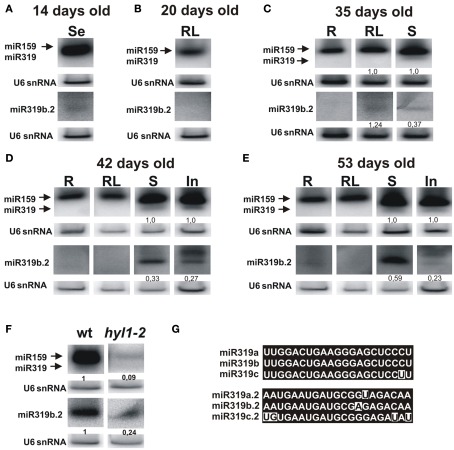
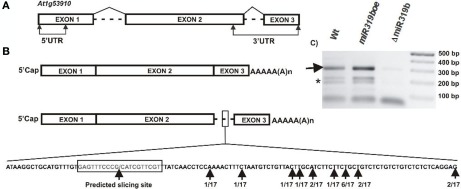
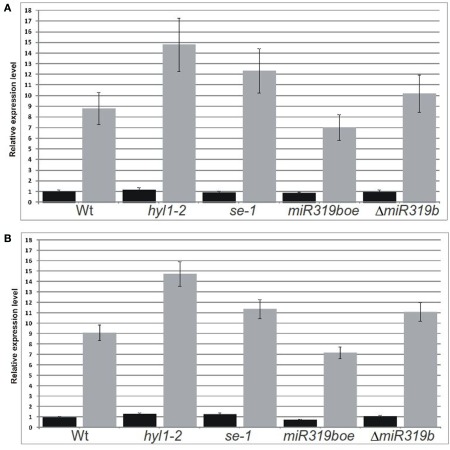
Arabidopsis miR319a/b/c primary transcripts are unusual due to the presence of a long stem and loop structure containing functional miR319a/b/c molecules. In our experiments carried out using high throughput sequencing (HTS), we have shown that additional microRNAs (miRNAs), miR319a.2/b.2/c.2 are generated from the upper part of the same hairpin structure. We have also found cognate miRNAa.2*/b.2*/c.2* to be present in the HTS results with a considerably lower number of reads. Northern hybridization revealed that miR319b.2 is mainly expressed in 35-day-old plant rosette leaves, as well as in stem and inflorescences of 42- and 53-day-old plants. Moreover, it carries multiple signatures of a functional miRNA, including as follows: (i) its biogenesis is HYL1-dependent; (ii) it is incorporated in a substantial amount into RISC complexes containing AGO1, AGO2, or AGO4 protein; (iii) 24 nt-long species of miR319b.2 have been found in inflorescences to be more abundant than 21 nt miR319b.2 species; (iv) it is present in various ratios to miR319b during plant development, which suggests the existence of a regulatory mechanism responsible for its biogenesis/processing; (v) there is an observed cross-species conservation of the miR319a/b/c stem nucleotide sequence extending beyond mature miRNA region; and (vi) all evidence suggests that intron-containing RAP2.12 mRNA isoform is the target for miR319b.2. All these features prompt us to claim miR319b.2 as a functional miRNA molecule.
Keywords: RAP2.12; gene expression; miRNA; pri-miRNA; splicing.
Figures

Similar articles
-
Arabidopsis microRNA expression regulation in a wide range of abiotic stress responses.Front Plant Sci. 2015 Jun 4;6:410. doi: 10.3389/fpls.2015.00410. eCollection 2015. Front Plant Sci. 2015. PMID: 26089831 Free PMC article.
-
Gene structures and processing of Arabidopsis thaliana HYL1-dependent pri-miRNAs.Nucleic Acids Res. 2009 May;37(9):3083-93. doi: 10.1093/nar/gkp189. Epub 2009 Mar 20. Nucleic Acids Res. 2009. PMID: 19304749 Free PMC article.
-
miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis.Proc Natl Acad Sci U S A. 2009 Dec 29;106(52):22534-9. doi: 10.1073/pnas.0908718106. Epub 2009 Dec 10. Proc Natl Acad Sci U S A. 2009. PMID: 20007771 Free PMC article.
-
Posttranscriptional coordination of splicing and miRNA biogenesis in plants.Wiley Interdiscip Rev RNA. 2017 May;8(3). doi: 10.1002/wrna.1403. Epub 2016 Nov 9. Wiley Interdiscip Rev RNA. 2017. PMID: 27863087 Review.
-
[Plant micro RNA biogenesis].Postepy Biochem. 2008;54(3):308-16. Postepy Biochem. 2008. PMID: 19112830 Review. Polish.
Cited by
-
Artificial microRNA guide strand selection from duplexes with no mismatches shows a purine-rich preference for virus- and non-virus-based expression vectors in plants.Plant Biotechnol J. 2022 Jun;20(6):1069-1084. doi: 10.1111/pbi.13786. Epub 2022 Feb 19. Plant Biotechnol J. 2022. PMID: 35113475 Free PMC article.
-
High-throughput sequencing and differential expression analysis of miRNAs in response to Brassinosteroid treatment in Arabidopsis thaliana.Funct Integr Genomics. 2019 Jul;19(4):597-615. doi: 10.1007/s10142-019-00668-1. Epub 2019 Feb 19. Funct Integr Genomics. 2019. PMID: 30783808
-
Regulatory non-coding RNAs: a new frontier in regulation of plant biology.Funct Integr Genomics. 2021 Jul;21(3-4):313-330. doi: 10.1007/s10142-021-00787-8. Epub 2021 May 20. Funct Integr Genomics. 2021. PMID: 34013486 Free PMC article. Review.
-
Alternative processing of its precursor is related to miR319 decreasing in melon plants exposed to cold.Sci Rep. 2018 Oct 19;8(1):15538. doi: 10.1038/s41598-018-34012-7. Sci Rep. 2018. PMID: 30341377 Free PMC article.
-
microRNA biogenesis and turnover in plants.Cold Spring Harb Symp Quant Biol. 2012;77:183-94. doi: 10.1101/sqb.2013.77.014530. Epub 2013 Feb 25. Cold Spring Harb Symp Quant Biol. 2012. PMID: 23439913 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials