

Review
doi: 10.1016/j.freeradbiomed.2012.05.008.
Epub 2012 May 15.
What really happens in the neutrophil phagosome?
Affiliations
- PMID: 22609248
- PMCID: PMC4382085
- DOI: 10.1016/j.freeradbiomed.2012.05.008
Item in Clipboard
Review
What really happens in the neutrophil phagosome?
Free Radic Biol Med.
.
Abstract
Current viewpoints concerning the bactericidal mechanisms of neutrophils are reviewed from a perspective that emphasizes challenges presented by the inability to duplicate ex vivo the intracellular milieu. Among the challenges considered are the influences of confinement upon substrate availability and reaction dynamics, direct and indirect synergistic interactions between individual toxins, and bacterial responses to stressors. Approaches to gauging relative contributions of various oxidative and nonoxidative toxins within neutrophils using bacteria and bacterial mimics as intrinsic probes are also discussed.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures

Four models of bactericidal action in neutrophils. Panel a: MPO-dependent oxidative killing. Granule-derived MPO catalyzes formation of HOCl from endogenous chloride and NOX-2-derived superoxide or hydrogen peroxide. HOCl and/or secondary chloramines formed by reaction with endogenous amines inactivate plasma membrane-localized proteins involved with energy transduction and biosynthesis (represented here as the F0F1-ATP synthase and a generic proton-symporting metabolite transport protein). Additional reactions of HOCl may involve formation of nitryl chloride (NO2Cl) if sufficient nitrite ion is present in the phagosomal fluid, and generation of hydroxyl and carbonate radicals following one-electron reduction by superoxide. Panel b: MPO-independent oxidative killing. Superoxide ion perfuses the bacterial membrane to react with iron-sulfur clusters in cytosolic dehydratases, releasing ferrous ion, which relocates to vulnerable target sites (here shown as genomic DNA). Subsequent site-specific oxidation by H2O2 in Fenton reactions leads to irreversible loss of function (here double-stranded cleavage of the DNA). (After Imlay and coworkers [53].) Additional (but presently controversial) pathways may involve singlet oxygen-initiated formation of hydrogen sesquioxide or ozone in antibody- or amino acid-catalyzed reactions. Panel c: Nonoxidative killing. Granule-derived cationic antimicrobial peptides (CAMP) aggregate in the bacterial plasma membranes to form membrane-spanning pores that dissipate ion gradients essential to homeostasis and energy transduction. In Gram-negative bacteria, granule-derived bactericidal permeability-increasing protein (BPI) binds to lipopolysaccharide (LPS), initiating a set of transformations that promote phospholipase (PLase) activation and access to and degradation of the bacterial membranes by these and other lytic proteins, including serum-derived complement (C7-C9) and group IIA-phospholipase A2 (PLA2). The latter gain access to the phagosome by binding to extracellular bacteria prior to phagocytosis. PG = peptidoglycan. (After Elsbach and coworkers [23,155].) Panel d: Redox-driven nonoxidative killing. Electrogenic NOX-2 respiration acts to polarize the phagosomal membrane, driving influx of electrolyte cations (here, K+). Hydrogen peroxide, formed as the respiratory end-product, is destroyed in catalatic reactions (here via MPO catalysis). The increased ionic strength causes release of granule-derived cationic seprocidins (e.g., cathepsin G, elastase) from anionic biopolymers, which then attack the bacterium. (After Segal and coworkers [41].)
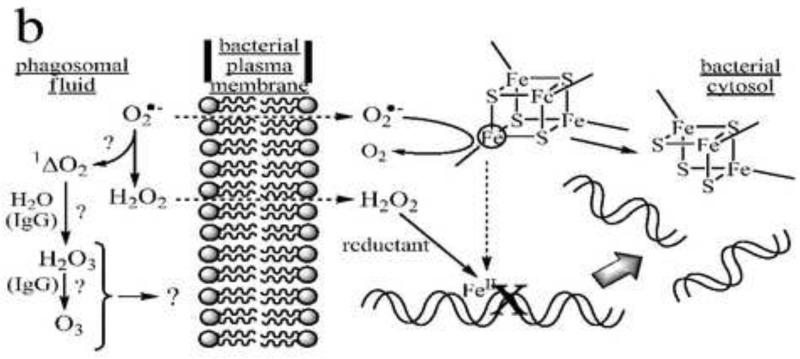
Four models of bactericidal action in neutrophils. Panel a: MPO-dependent oxidative killing. Granule-derived MPO catalyzes formation of HOCl from endogenous chloride and NOX-2-derived superoxide or hydrogen peroxide. HOCl and/or secondary chloramines formed by reaction with endogenous amines inactivate plasma membrane-localized proteins involved with energy transduction and biosynthesis (represented here as the F0F1-ATP synthase and a generic proton-symporting metabolite transport protein). Additional reactions of HOCl may involve formation of nitryl chloride (NO2Cl) if sufficient nitrite ion is present in the phagosomal fluid, and generation of hydroxyl and carbonate radicals following one-electron reduction by superoxide. Panel b: MPO-independent oxidative killing. Superoxide ion perfuses the bacterial membrane to react with iron-sulfur clusters in cytosolic dehydratases, releasing ferrous ion, which relocates to vulnerable target sites (here shown as genomic DNA). Subsequent site-specific oxidation by H2O2 in Fenton reactions leads to irreversible loss of function (here double-stranded cleavage of the DNA). (After Imlay and coworkers [53].) Additional (but presently controversial) pathways may involve singlet oxygen-initiated formation of hydrogen sesquioxide or ozone in antibody- or amino acid-catalyzed reactions. Panel c: Nonoxidative killing. Granule-derived cationic antimicrobial peptides (CAMP) aggregate in the bacterial plasma membranes to form membrane-spanning pores that dissipate ion gradients essential to homeostasis and energy transduction. In Gram-negative bacteria, granule-derived bactericidal permeability-increasing protein (BPI) binds to lipopolysaccharide (LPS), initiating a set of transformations that promote phospholipase (PLase) activation and access to and degradation of the bacterial membranes by these and other lytic proteins, including serum-derived complement (C7-C9) and group IIA-phospholipase A2 (PLA2). The latter gain access to the phagosome by binding to extracellular bacteria prior to phagocytosis. PG = peptidoglycan. (After Elsbach and coworkers [23,155].) Panel d: Redox-driven nonoxidative killing. Electrogenic NOX-2 respiration acts to polarize the phagosomal membrane, driving influx of electrolyte cations (here, K+). Hydrogen peroxide, formed as the respiratory end-product, is destroyed in catalatic reactions (here via MPO catalysis). The increased ionic strength causes release of granule-derived cationic seprocidins (e.g., cathepsin G, elastase) from anionic biopolymers, which then attack the bacterium. (After Segal and coworkers [41].)

Four models of bactericidal action in neutrophils. Panel a: MPO-dependent oxidative killing. Granule-derived MPO catalyzes formation of HOCl from endogenous chloride and NOX-2-derived superoxide or hydrogen peroxide. HOCl and/or secondary chloramines formed by reaction with endogenous amines inactivate plasma membrane-localized proteins involved with energy transduction and biosynthesis (represented here as the F0F1-ATP synthase and a generic proton-symporting metabolite transport protein). Additional reactions of HOCl may involve formation of nitryl chloride (NO2Cl) if sufficient nitrite ion is present in the phagosomal fluid, and generation of hydroxyl and carbonate radicals following one-electron reduction by superoxide. Panel b: MPO-independent oxidative killing. Superoxide ion perfuses the bacterial membrane to react with iron-sulfur clusters in cytosolic dehydratases, releasing ferrous ion, which relocates to vulnerable target sites (here shown as genomic DNA). Subsequent site-specific oxidation by H2O2 in Fenton reactions leads to irreversible loss of function (here double-stranded cleavage of the DNA). (After Imlay and coworkers [53].) Additional (but presently controversial) pathways may involve singlet oxygen-initiated formation of hydrogen sesquioxide or ozone in antibody- or amino acid-catalyzed reactions. Panel c: Nonoxidative killing. Granule-derived cationic antimicrobial peptides (CAMP) aggregate in the bacterial plasma membranes to form membrane-spanning pores that dissipate ion gradients essential to homeostasis and energy transduction. In Gram-negative bacteria, granule-derived bactericidal permeability-increasing protein (BPI) binds to lipopolysaccharide (LPS), initiating a set of transformations that promote phospholipase (PLase) activation and access to and degradation of the bacterial membranes by these and other lytic proteins, including serum-derived complement (C7-C9) and group IIA-phospholipase A2 (PLA2). The latter gain access to the phagosome by binding to extracellular bacteria prior to phagocytosis. PG = peptidoglycan. (After Elsbach and coworkers [23,155].) Panel d: Redox-driven nonoxidative killing. Electrogenic NOX-2 respiration acts to polarize the phagosomal membrane, driving influx of electrolyte cations (here, K+). Hydrogen peroxide, formed as the respiratory end-product, is destroyed in catalatic reactions (here via MPO catalysis). The increased ionic strength causes release of granule-derived cationic seprocidins (e.g., cathepsin G, elastase) from anionic biopolymers, which then attack the bacterium. (After Segal and coworkers [41].)

Four models of bactericidal action in neutrophils. Panel a: MPO-dependent oxidative killing. Granule-derived MPO catalyzes formation of HOCl from endogenous chloride and NOX-2-derived superoxide or hydrogen peroxide. HOCl and/or secondary chloramines formed by reaction with endogenous amines inactivate plasma membrane-localized proteins involved with energy transduction and biosynthesis (represented here as the F0F1-ATP synthase and a generic proton-symporting metabolite transport protein). Additional reactions of HOCl may involve formation of nitryl chloride (NO2Cl) if sufficient nitrite ion is present in the phagosomal fluid, and generation of hydroxyl and carbonate radicals following one-electron reduction by superoxide. Panel b: MPO-independent oxidative killing. Superoxide ion perfuses the bacterial membrane to react with iron-sulfur clusters in cytosolic dehydratases, releasing ferrous ion, which relocates to vulnerable target sites (here shown as genomic DNA). Subsequent site-specific oxidation by H2O2 in Fenton reactions leads to irreversible loss of function (here double-stranded cleavage of the DNA). (After Imlay and coworkers [53].) Additional (but presently controversial) pathways may involve singlet oxygen-initiated formation of hydrogen sesquioxide or ozone in antibody- or amino acid-catalyzed reactions. Panel c: Nonoxidative killing. Granule-derived cationic antimicrobial peptides (CAMP) aggregate in the bacterial plasma membranes to form membrane-spanning pores that dissipate ion gradients essential to homeostasis and energy transduction. In Gram-negative bacteria, granule-derived bactericidal permeability-increasing protein (BPI) binds to lipopolysaccharide (LPS), initiating a set of transformations that promote phospholipase (PLase) activation and access to and degradation of the bacterial membranes by these and other lytic proteins, including serum-derived complement (C7-C9) and group IIA-phospholipase A2 (PLA2). The latter gain access to the phagosome by binding to extracellular bacteria prior to phagocytosis. PG = peptidoglycan. (After Elsbach and coworkers [23,155].) Panel d: Redox-driven nonoxidative killing. Electrogenic NOX-2 respiration acts to polarize the phagosomal membrane, driving influx of electrolyte cations (here, K+). Hydrogen peroxide, formed as the respiratory end-product, is destroyed in catalatic reactions (here via MPO catalysis). The increased ionic strength causes release of granule-derived cationic seprocidins (e.g., cathepsin G, elastase) from anionic biopolymers, which then attack the bacterium. (After Segal and coworkers [41].)
Similar articles
-
Epic Immune Battles of History: Neutrophils vs. Staphylococcus aureus.Front Cell Infect Microbiol. 2017 Jun 30;7:286. doi: 10.3389/fcimb.2017.00286. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28713774 Free PMC article. Review.
-
Filifactor alocis modulates human neutrophil antimicrobial functional responses.Cell Microbiol. 2018 Jun;20(6):e12829. doi: 10.1111/cmi.12829. Epub 2018 Feb 12. Cell Microbiol. 2018. PMID: 29377528 Free PMC article.
-
Immunology. Lethal weapons.Science. 2002 Apr 26;296(5568):669-71. doi: 10.1126/science.1071271. Science. 2002. PMID: 11976433 No abstract available.
-
Analysis of Neutrophil Bactericidal Activity.Methods Mol Biol. 2020;2087:149-164. doi: 10.1007/978-1-0716-0154-9_12. Methods Mol Biol. 2020. PMID: 31728990
-
Staphylococcus aureus host cell invasion and post-invasion events.Int J Med Microbiol. 2010 Feb;300(2-3):170-5. doi: 10.1016/j.ijmm.2009.08.019. Epub 2009 Sep 24. Int J Med Microbiol. 2010. PMID: 19781990 Review.
Cited by
-
Host Cell Antimicrobial Responses against Helicobacter pylori Infection: From Biological Aspects to Therapeutic Strategies.Int J Mol Sci. 2022 Sep 19;23(18):10941. doi: 10.3390/ijms231810941. Int J Mol Sci. 2022. PMID: 36142852 Free PMC article. Review.
-
Catastrophic chromosome fragmentation probes the nucleoid structure and dynamics in Escherichia coli.Nucleic Acids Res. 2022 Oct 28;50(19):11013-11027. doi: 10.1093/nar/gkac865. Nucleic Acids Res. 2022. PMID: 36243965 Free PMC article.
-
Sequential action of antibacterial effectors in Dictyostelium discoideum phagosomes.Mol Microbiol. 2023 Jan;119(1):74-85. doi: 10.1111/mmi.15004. Epub 2022 Dec 4. Mol Microbiol. 2023. PMID: 36416195 Free PMC article.
-
Mechanism of Hip Arthropathy in Ankylosing Spondylitis: Abnormal Myeloperoxidase and Phagosome.Front Immunol. 2021 Nov 22;12:572592. doi: 10.3389/fimmu.2021.572592. eCollection 2021. Front Immunol. 2021. PMID: 34880852 Free PMC article.
-
Modeling Host-Pathogen Interaction to Elucidate the Metabolic Drug Response of Intracellular Mycobacterium tuberculosis.Front Cell Infect Microbiol. 2019 May 8;9:144. doi: 10.3389/fcimb.2019.00144. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31139575 Free PMC article.
References
-
- DeLeo FR, Allen L-AH, Apicella M, Nauseef WM. NADPH oxidase activation and assembly during phagocytosis. J. Immunol. 1999;163:6732–6740. - PubMed
-
- Jiang Q, Griffin DA, Barofsky DA, Hurst JK. Intraphagosomal chlorination dynamics and yields determined using unique fluorescent bacterial mimics. Chem. Res. Toxicol. 1997;10:1080–1089. - PubMed
-
- Pan L, Zhang X, Song K, Tang B, Cai W, Wu X, Rupp RA, Xu J. Real-time imaging of autofluorescence NAD(P)H in single human neutrophils. Appl. Optics. 2009;48:1042–1046. - PubMed
-
- Ohno Y-I, Hirai K-I, Kanoh T, Uchino H, Ogawa K. Subcellular localization of H2O2 production in human neutrophils stimulated with particles and an effect of cytochalasin B on the cells. Blood. 1982;60:253–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
