

Lack of Cul4b, an E3 ubiquitin ligase component, leads to embryonic lethality and abnormal placental development
- PMID: 22606329
- PMCID: PMC3351389
- DOI: 10.1371/journal.pone.0037070
Lack of Cul4b, an E3 ubiquitin ligase component, leads to embryonic lethality and abnormal placental development
Abstract
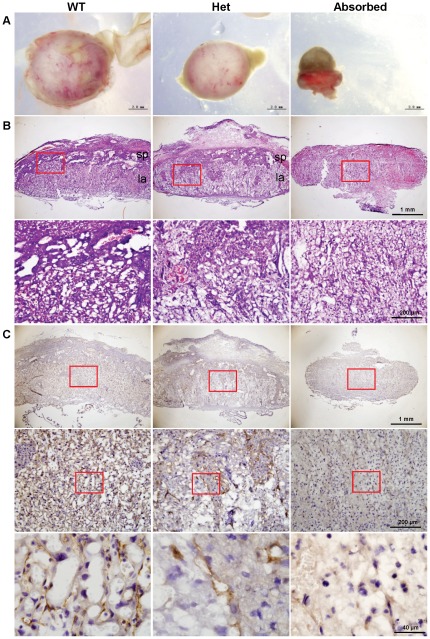
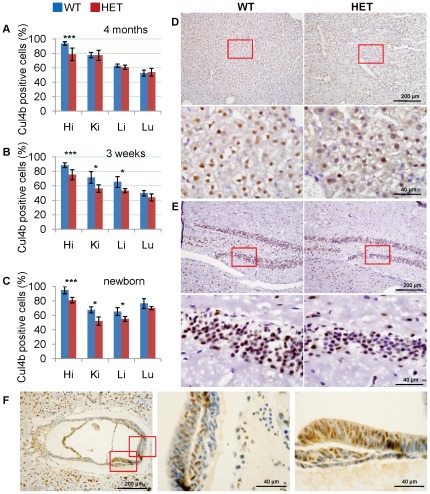
Cullin-RING ligases (CRLs) complexes participate in the regulation of diverse cellular processes, including cell cycle progression, transcription, signal transduction and development. Serving as the scaffold protein, cullins are crucial for the assembly of ligase complexes, which recognize and target various substrates for proteosomal degradation. Mutations in human CUL4B, one of the eight members in cullin family, are one of the major causes of X-linked mental retardation. We here report the generation and characterization of Cul4b knockout mice, in which exons 3 to 5 were deleted. In contrast to the survival to adulthood of human hemizygous males with CUL4B null mutation, Cul4b null mouse embryos show severe developmental arrest and usually die before embryonic day 9.5 (E9.5). Accumulation of cyclin E, a CRL (CUL4B) substrate, was observed in Cul4b null embryos. Cul4b heterozygotes were recovered at a reduced ratio and exhibited a severe developmental delay. The placentas in Cul4b heterozygotes were disorganized and were impaired in vascularization, which may contribute to the developmental delay. As in human CUL4B heterozygotes, Cul4b null cells were selected against in Cul4b heterozygotes, leading to various degrees of skewed X-inactivation in different tissues. Together, our results showed that CUL4B is indispensable for embryonic development in the mouse.
Conflict of interest statement
Figures
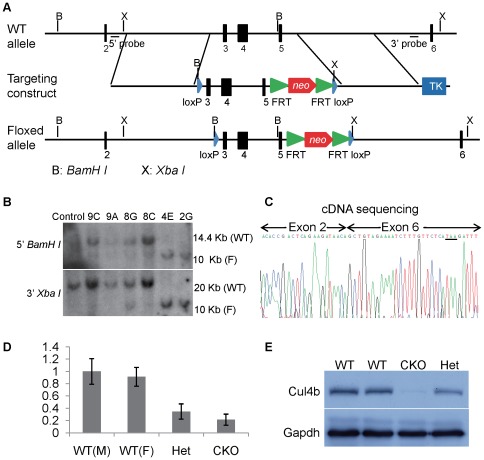
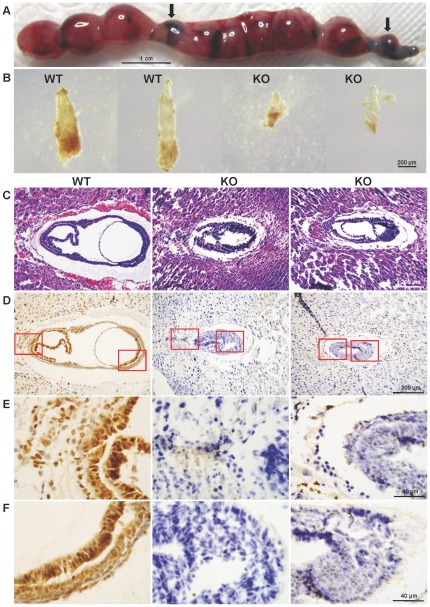
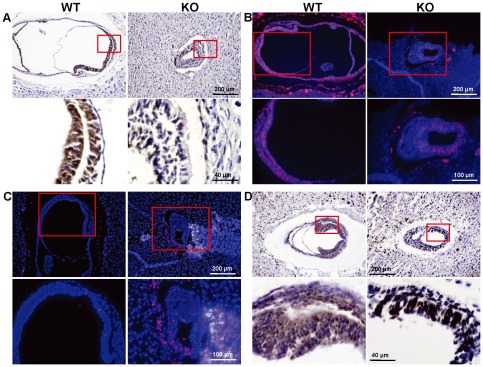
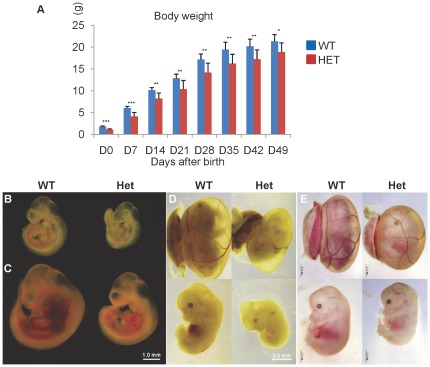
Similar articles
-
Essential role of the CUL4B ubiquitin ligase in extra-embryonic tissue development during mouse embryogenesis.Cell Res. 2012 Aug;22(8):1258-69. doi: 10.1038/cr.2012.48. Epub 2012 Mar 27. Cell Res. 2012. PMID: 22453236 Free PMC article.
-
CUL4B ubiquitin ligase in mouse development: a model for human X-linked mental retardation syndrome?Cell Res. 2012 Aug;22(8):1224-6. doi: 10.1038/cr.2012.79. Epub 2012 May 15. Cell Res. 2012. PMID: 22584258 Free PMC article.
-
Rescue of the genetically engineered Cul4b mutant mouse as a potential model for human X-linked mental retardation.Hum Mol Genet. 2012 Oct 1;21(19):4270-85. doi: 10.1093/hmg/dds261. Epub 2012 Jul 3. Hum Mol Genet. 2012. PMID: 22763239
-
Distinct and overlapping functions of the cullin E3 ligase scaffolding proteins CUL4A and CUL4B.Gene. 2015 Nov 15;573(1):33-45. doi: 10.1016/j.gene.2015.08.064. Epub 2015 Sep 3. Gene. 2015. PMID: 26344709 Free PMC article. Review.
-
CUL4B-deficiency in humans: understanding the clinical consequences of impaired Cullin 4-RING E3 ubiquitin ligase function.Mech Ageing Dev. 2011 Aug;132(8-9):366-73. doi: 10.1016/j.mad.2011.02.003. Epub 2011 Feb 23. Mech Ageing Dev. 2011. PMID: 21352845 Review.
Cited by
-
Genetically engineered mouse models for functional studies of SKP1-CUL1-F-box-protein (SCF) E3 ubiquitin ligases.Cell Res. 2013 May;23(5):599-619. doi: 10.1038/cr.2013.44. Epub 2013 Mar 26. Cell Res. 2013. PMID: 23528706 Free PMC article. Review.
-
Accelerated hepatocellular carcinoma development in CUL4B transgenic mice.Oncotarget. 2015 Jun 20;6(17):15209-21. doi: 10.18632/oncotarget.3829. Oncotarget. 2015. PMID: 25945838 Free PMC article.
-
Cullin-RING E3 ubiquitin ligase 4 regulates neurite morphogenesis during neurodevelopment.iScience. 2024 Jan 17;27(2):108933. doi: 10.1016/j.isci.2024.108933. eCollection 2024 Feb 16. iScience. 2024. PMID: 38318354 Free PMC article.
-
CUL4B functions as a tumor suppressor in KRAS-driven lung tumors by inhibiting the recruitment of myeloid-derived suppressor cells.Oncogene. 2023 Oct;42(42):3113-3126. doi: 10.1038/s41388-023-02824-1. Epub 2023 Aug 31. Oncogene. 2023. PMID: 37653114
-
PHD finger protein 1 (PHF1) is a novel reader for histone H4R3 symmetric dimethylation and coordinates with PRMT5-WDR77/CRL4B complex to promote tumorigenesis.Nucleic Acids Res. 2018 Jul 27;46(13):6608-6626. doi: 10.1093/nar/gky461. Nucleic Acids Res. 2018. PMID: 29846670 Free PMC article.
References
-
- Petroski MD, Deshaies RJ. Function and regulation of cullin-RING ubiquitin ligases. Nat Rev Mol Cell Biol. 2005;6:9–20. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
