

Regulation of ribosomal RNA production by RNA polymerase I: does elongation come first?
- PMID: 22567380
- PMCID: PMC3335655
- DOI: 10.1155/2012/276948
Regulation of ribosomal RNA production by RNA polymerase I: does elongation come first?
Abstract
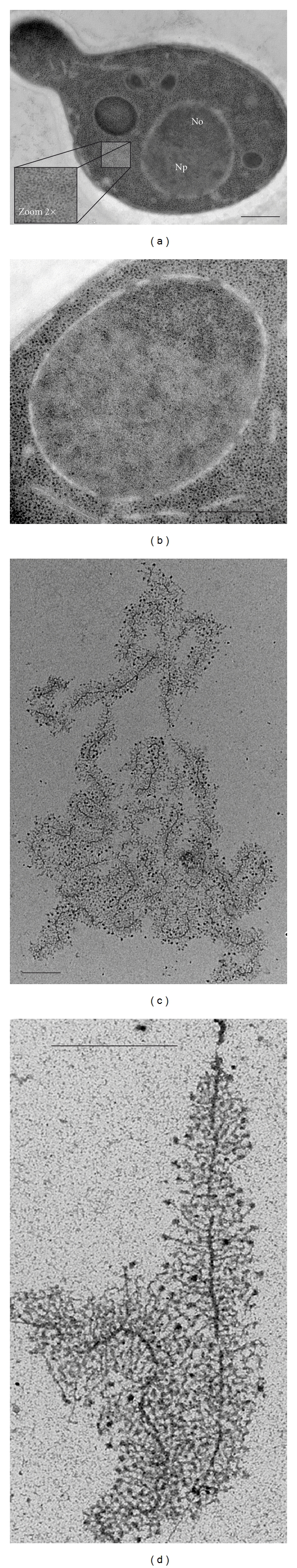
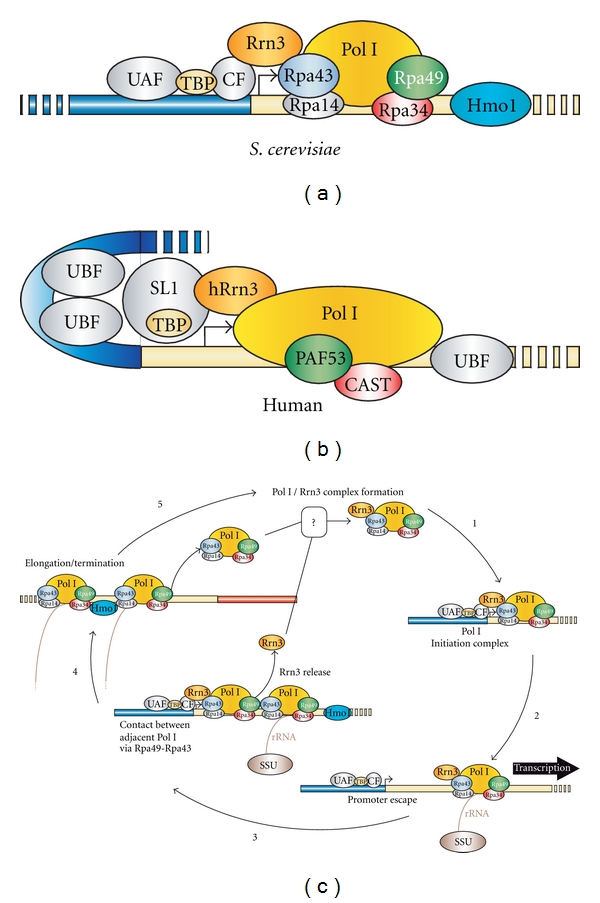
Ribosomal RNA (rRNA) production represents the most active transcription in the cell. Synthesis of the large rRNA precursors (35-47S) can be achieved by up to 150 RNA polymerase I (Pol I) enzymes simultaneously transcribing each rRNA gene. In this paper, we present recent advances made in understanding the regulatory mechanisms that control elongation. Built-in Pol I elongation factors, such as Rpa34/Rpa49 in budding yeast and PAF53/CAST in humans, are instrumental to the extremely high rate of rRNA production per gene. rRNA elongation mechanisms are intrinsically linked to chromatin structure and to the higher-order organization of the rRNA genes (rDNA). Factors such as Hmo1 in yeast and UBF1 in humans are key players in rDNA chromatin structure in vivo. Finally, elongation factors known to regulate messengers RNA production by RNA polymerase II are also involved in rRNA production and work cooperatively with Rpa49 in vivo.
Figures

Similar articles
-
Genetic analyses led to the discovery of a super-active mutant of the RNA polymerase I.PLoS Genet. 2019 May 28;15(5):e1008157. doi: 10.1371/journal.pgen.1008157. eCollection 2019 May. PLoS Genet. 2019. PMID: 31136569 Free PMC article.
-
Coupling Between Production of Ribosomal RNA and Maturation: Just at the Beginning.Front Mol Biosci. 2021 Oct 26;8:778778. doi: 10.3389/fmolb.2021.778778. eCollection 2021. Front Mol Biosci. 2021. PMID: 34765647 Free PMC article. Review.
-
RNA polymerase I (Pol I) passage through nucleosomes depends on Pol I subunits binding its lobe structure.J Biol Chem. 2020 Apr 10;295(15):4782-4795. doi: 10.1074/jbc.RA119.011827. Epub 2020 Feb 14. J Biol Chem. 2020. PMID: 32060094 Free PMC article.
-
Two RNA polymerase I subunits control the binding and release of Rrn3 during transcription.Mol Cell Biol. 2008 Mar;28(5):1596-605. doi: 10.1128/MCB.01464-07. Epub 2007 Dec 17. Mol Cell Biol. 2008. PMID: 18086878 Free PMC article.
-
Coordinated Control of rRNA Processing by RNA Polymerase I.Trends Genet. 2019 Oct;35(10):724-733. doi: 10.1016/j.tig.2019.07.002. Epub 2019 Jul 26. Trends Genet. 2019. PMID: 31358304 Free PMC article. Review.
Cited by
-
Compositional reorganization of the nucleolus in budding yeast mitosis.Mol Biol Cell. 2019 Mar 1;30(5):591-606. doi: 10.1091/mbc.E18-08-0524. Epub 2019 Jan 9. Mol Biol Cell. 2019. PMID: 30625028 Free PMC article.
-
Impaired ribosome biogenesis: mechanisms and relevance to cancer and aging.Aging (Albany NY). 2019 Apr 26;11(8):2512-2540. doi: 10.18632/aging.101922. Aging (Albany NY). 2019. PMID: 31026227 Free PMC article. Review.
-
Yeast HMO1: Linker Histone Reinvented.Microbiol Mol Biol Rev. 2016 Nov 30;81(1):e00037-16. doi: 10.1128/MMBR.00037-16. Print 2017 Mar. Microbiol Mol Biol Rev. 2016. PMID: 27903656 Free PMC article. Review.
-
A dynamic ribosomal biogenesis response is not required for IGF-1-mediated hypertrophy of human primary myotubes.FASEB J. 2017 Dec;31(12):5196-5207. doi: 10.1096/fj.201700329R. Epub 2017 Aug 3. FASEB J. 2017. PMID: 28774889 Free PMC article.
-
Glutamine methylation in histone H2A is an RNA-polymerase-I-dedicated modification.Nature. 2014 Jan 23;505(7484):564-8. doi: 10.1038/nature12819. Epub 2013 Dec 18. Nature. 2014. PMID: 24352239 Free PMC article.
References
-
- Warner JR. The economics of ribosome biosynthesis in yeast. Trends in Biochemical Sciences. 1999;24(11):437–440. - PubMed
-
- Miller OL, Beatty BR. Visualization of nucleolar genes. Science. 1969;164(3882):955–957. - PubMed
-
- Schweizer E, Halvorson HO. On the regulation of ribosomal RNA synthesis in yeast. Experimental Cell Research. 1969;56(2-3):239–244. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
