

Monomeric site-specific nucleases for genome editing
- PMID: 22566637
- PMCID: PMC3361397
- DOI: 10.1073/pnas.1117984109
Monomeric site-specific nucleases for genome editing
Abstract
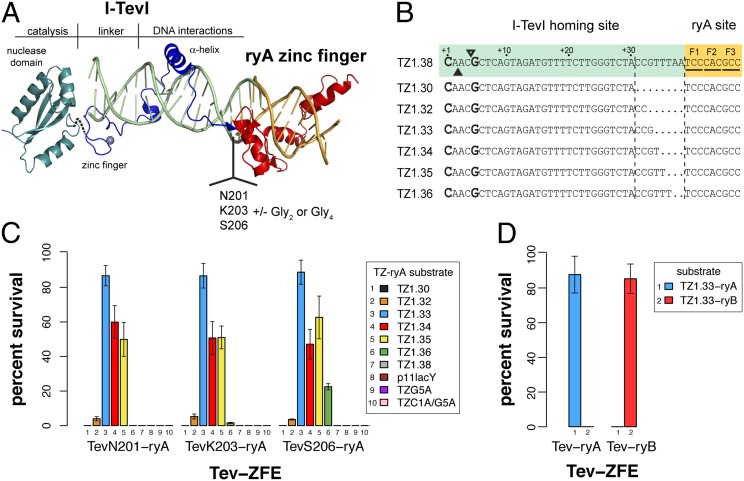
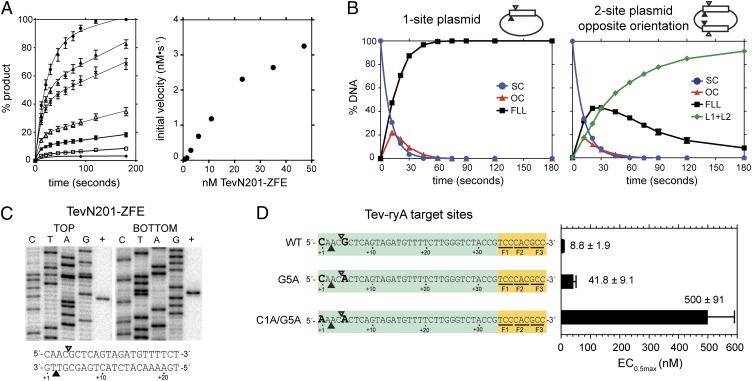
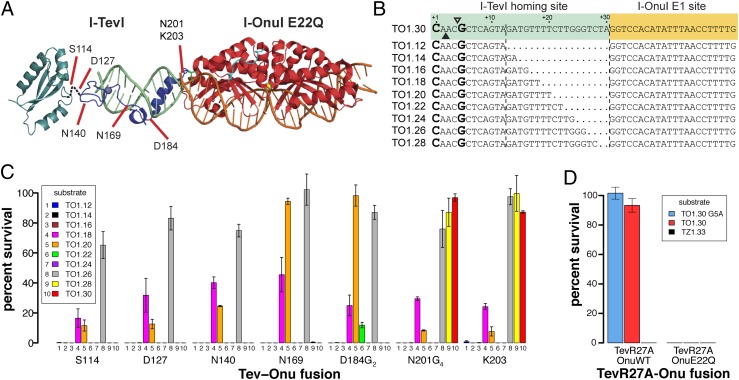
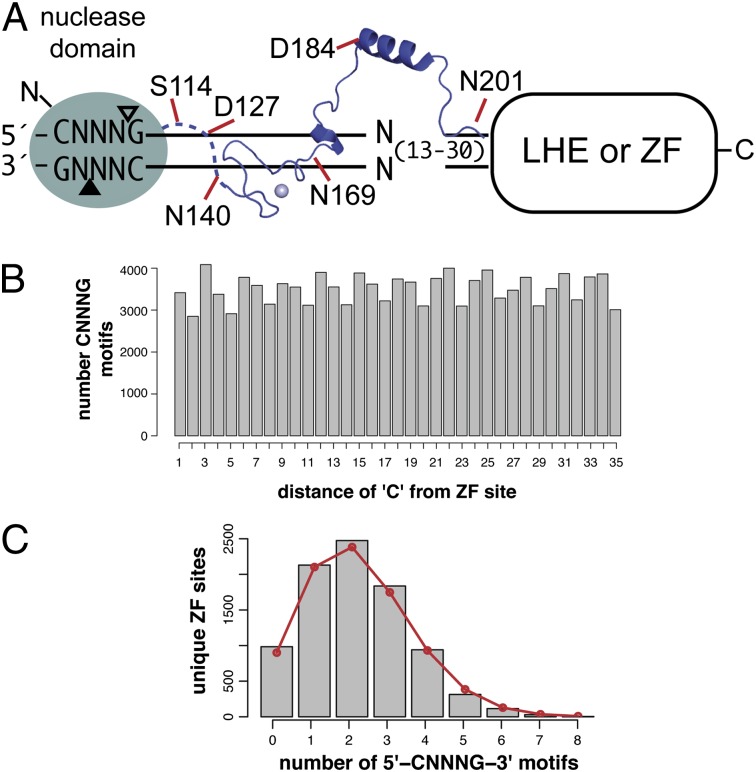
Targeted manipulation of complex genomes often requires the introduction of a double-strand break at defined locations by site-specific DNA endonucleases. Here, we describe a monomeric nuclease domain derived from GIY-YIG homing endonucleases for genome-editing applications. Fusion of the GIY-YIG nuclease domain to three-member zinc-finger DNA binding domains generated chimeric GIY-zinc finger endonucleases (GIY-ZFEs). Significantly, the I-TevI-derived fusions (Tev-ZFEs) function in vitro as monomers to introduce a double-strand break, and discriminate in vitro and in bacterial and yeast assays against substrates lacking a preferred 5'-CNNNG-3' cleavage motif. The Tev-ZFEs function to induce recombination in a yeast-based assay with activity on par with a homodimeric Zif268 zinc-finger nuclease. We also fused the I-TevI nuclease domain to a catalytically inactive LADGLIDADG homing endonuclease (LHE) scaffold. The monomeric Tev-LHEs are active in vivo and similarly discriminate against substrates lacking the 5'-CNNNG-3' motif. The monomeric Tev-ZFEs and Tev-LHEs are distinct from the FokI-derived zinc-finger nuclease and TAL effector nuclease platforms as the GIY-YIG domain alleviates the requirement to design two nuclease fusions to target a given sequence, highlighting the diversity of nuclease domains with distinctive biochemical properties suitable for genome-editing applications.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
The monomeric GIY-YIG homing endonuclease I-BmoI uses a molecular anchor and a flexible tether to sequentially nick DNA.Nucleic Acids Res. 2013 May 1;41(10):5413-27. doi: 10.1093/nar/gkt186. Epub 2013 Apr 4. Nucleic Acids Res. 2013. PMID: 23558745 Free PMC article.
-
The I-TevI nuclease and linker domains contribute to the specificity of monomeric TALENs.G3 (Bethesda). 2014 Apr 16;4(6):1155-65. doi: 10.1534/g3.114.011445. G3 (Bethesda). 2014. PMID: 24739648 Free PMC article.
-
Oligomeric structure diversity within the GIY-YIG nuclease family.J Mol Biol. 2009 Mar 20;387(1):10-6. doi: 10.1016/j.jmb.2009.01.048. Epub 2009 Jan 30. J Mol Biol. 2009. PMID: 19361436
-
Applications of Alternative Nucleases in the Age of CRISPR/Cas9.Int J Mol Sci. 2017 Nov 29;18(12):2565. doi: 10.3390/ijms18122565. Int J Mol Sci. 2017. PMID: 29186020 Free PMC article. Review.
-
The democratization of gene editing: Insights from site-specific cleavage and double-strand break repair.DNA Repair (Amst). 2016 Aug;44:6-16. doi: 10.1016/j.dnarep.2016.05.001. Epub 2016 May 12. DNA Repair (Amst). 2016. PMID: 27261202 Free PMC article. Review.
Cited by
-
To nick or not to nick: comparison of I-SceI single- and double-strand break-induced recombination in yeast and human cells.PLoS One. 2014 Feb 18;9(2):e88840. doi: 10.1371/journal.pone.0088840. eCollection 2014. PLoS One. 2014. PMID: 24558436 Free PMC article.
-
Homing endonucleases from mobile group I introns: discovery to genome engineering.Mob DNA. 2014 Mar 3;5(1):7. doi: 10.1186/1759-8753-5-7. Mob DNA. 2014. PMID: 24589358 Free PMC article.
-
Repair of single-point mutations by polypurine reverse Hoogsteen hairpins.Hum Gene Ther Methods. 2014 Oct;25(5):288-302. doi: 10.1089/hgtb.2014.049. Epub 2014 Oct 14. Hum Gene Ther Methods. 2014. PMID: 25222154 Free PMC article.
-
Cas9 as a versatile tool for engineering biology.Nat Methods. 2013 Oct;10(10):957-63. doi: 10.1038/nmeth.2649. Nat Methods. 2013. PMID: 24076990 Free PMC article. Review.
-
The monomeric GIY-YIG homing endonuclease I-BmoI uses a molecular anchor and a flexible tether to sequentially nick DNA.Nucleic Acids Res. 2013 May 1;41(10):5413-27. doi: 10.1093/nar/gkt186. Epub 2013 Apr 4. Nucleic Acids Res. 2013. PMID: 23558745 Free PMC article.
References
-
- Bibikova M, Beumer K, Trautman JK, Carroll D. Enhancing gene targeting with designed zinc finger nucleases. Science. 2003;300:764. - PubMed
-
- Bogdanove AJ, Voytas DF. TAL effectors: Customizable proteins for DNA targeting. Science. 2011;333:1843–1846. - PubMed
-
- Urnov FD, Rebar EJ, Holmes MC, Zhang HS, Gregory PD. Genome editing with engineered zinc finger nucleases. Nat Rev Genet. 2010;11:636–646. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
