

Mechanisms contributing to central excitability changes during hearing loss
- PMID: 22566618
- PMCID: PMC3361412
- DOI: 10.1073/pnas.1116981109
Mechanisms contributing to central excitability changes during hearing loss
Abstract
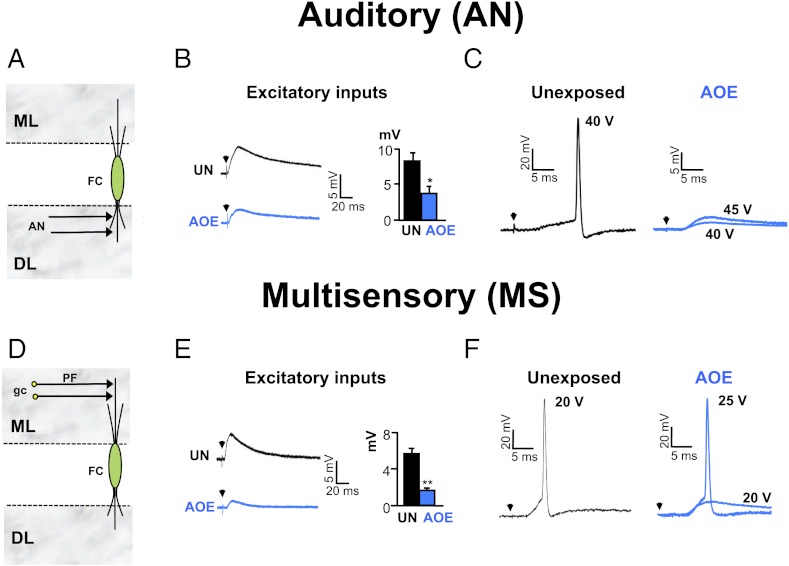
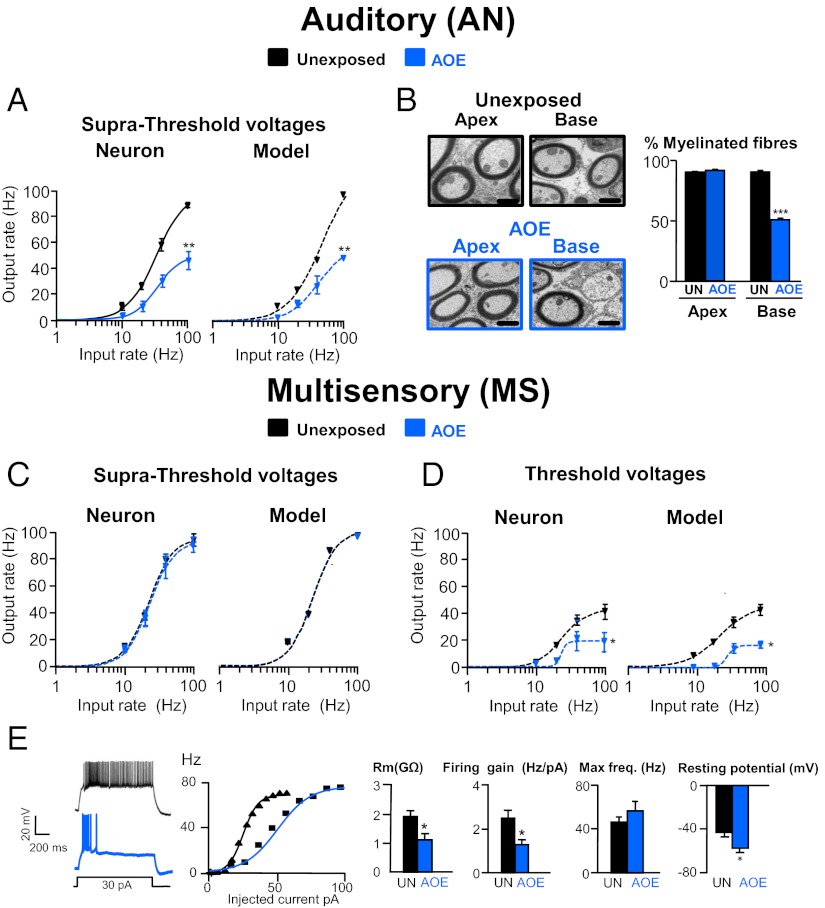
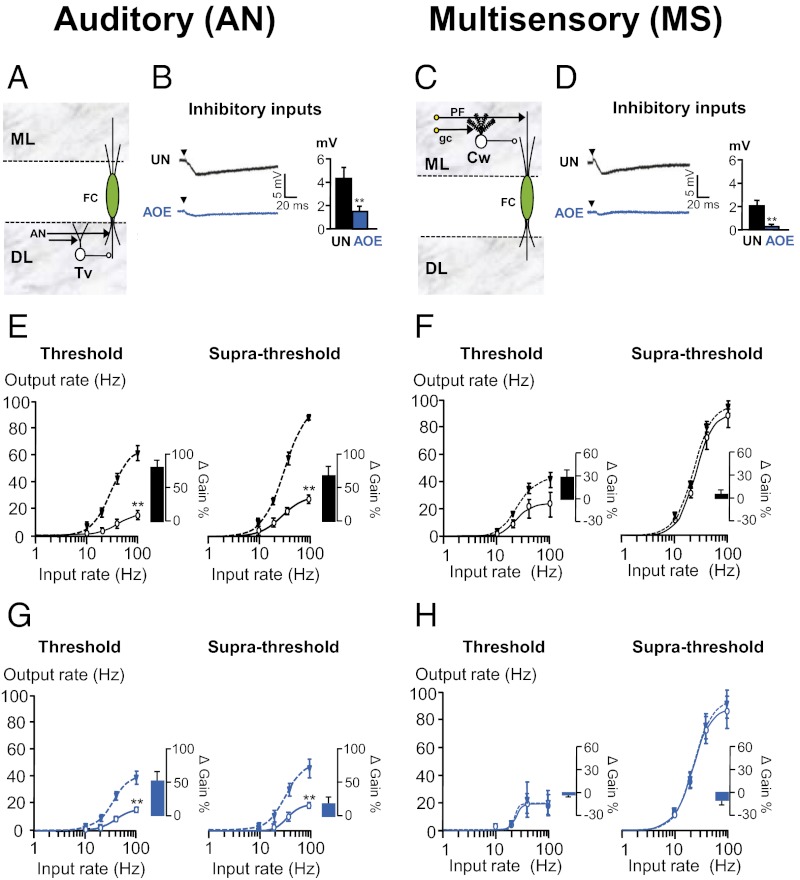
Exposure to loud sound causes cochlear damage resulting in hearing loss and tinnitus. Tinnitus has been related to hyperactivity in the central auditory pathway occurring weeks after loud sound exposure. However, central excitability changes concomitant to hearing loss and preceding those periods of hyperactivity, remain poorly explored. Here we investigate mechanisms contributing to excitability changes in the dorsal cochlear nucleus (DCN) shortly after exposure to loud sound that produces hearing loss. We show that acoustic overexposure alters synaptic transmission originating from the auditory and the multisensory pathway within the DCN in different ways. A reduction in the number of myelinated auditory nerve fibers leads to a reduced maximal firing rate of DCN principal cells, which cannot be restored by increasing auditory nerve fiber recruitment. In contrast, a decreased membrane resistance of DCN granule cells (multisensory inputs) leads to a reduced maximal firing rate of DCN principal cells that is overcome when additional multisensory fibers are recruited. Furthermore, gain modulation by inhibitory synaptic transmission is disabled in both auditory and multisensory pathways. These cellular mechanisms that contribute to decreased cellular excitability in the central auditory pathway are likely to represent early neurobiological markers of hearing loss and may suggest interventions to delay or stop the development of hyperactivity that has been associated with tinnitus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Development of hyperactivity after hearing loss in a computational model of the dorsal cochlear nucleus depends on neuron response type.Hear Res. 2008 Jun;240(1-2):57-72. doi: 10.1016/j.heares.2008.02.006. Epub 2008 Feb 26. Hear Res. 2008. PMID: 18396381
-
Acoustic over-exposure triggers burst firing in dorsal cochlear nucleus fusiform cells.Hear Res. 2012 Jan;283(1-2):98-106. doi: 10.1016/j.heares.2011.10.008. Epub 2011 Nov 7. Hear Res. 2012. PMID: 22085487 Free PMC article.
-
Dorsal cochlear nucleus responses to somatosensory stimulation are enhanced after noise-induced hearing loss.Eur J Neurosci. 2008 Jan;27(1):155-68. doi: 10.1111/j.1460-9568.2007.05983.x. Eur J Neurosci. 2008. PMID: 18184319 Free PMC article.
-
The dorsal cochlear nucleus as a contributor to tinnitus: mechanisms underlying the induction of hyperactivity.Prog Brain Res. 2007;166:89-106. doi: 10.1016/S0079-6123(07)66009-9. Prog Brain Res. 2007. PMID: 17956775 Review.
-
Dorsal cochlear nucleus hyperactivity and tinnitus: are they related?Am J Audiol. 2008 Dec;17(2):S148-61. doi: 10.1044/1059-0889(2008/08-0004). Epub 2008 Oct 31. Am J Audiol. 2008. PMID: 18978198 Review.
Cited by
-
Noise-Induced "Toughening" Effect in Wistar Rats: Enhanced Auditory Brainstem Responses Are Related to Calretinin and Nitric Oxide Synthase Upregulation.Front Neuroanat. 2016 Mar 31;10:19. doi: 10.3389/fnana.2016.00019. eCollection 2016. Front Neuroanat. 2016. PMID: 27065815 Free PMC article.
-
Alterations in regional homogeneity of resting-state brain activity in internet gaming addicts.Behav Brain Funct. 2012 Aug 18;8:41. doi: 10.1186/1744-9081-8-41. Behav Brain Funct. 2012. PMID: 22901705 Free PMC article.
-
Acoustic Trauma Changes the Parvalbumin-Positive Neurons in Rat Auditory Cortex.Neural Plast. 2018 Feb 8;2018:9828070. doi: 10.1155/2018/9828070. eCollection 2018. Neural Plast. 2018. PMID: 29593786 Free PMC article.
-
Map plasticity following noise exposure in auditory cortex of rats: implications for disentangling neural correlates of tinnitus and hyperacusis.Front Neurosci. 2024 May 31;18:1385942. doi: 10.3389/fnins.2024.1385942. eCollection 2024. Front Neurosci. 2024. PMID: 38881748 Free PMC article.
-
Enhanced GABAA-Mediated Tonic Inhibition in Auditory Thalamus of Rats with Behavioral Evidence of Tinnitus.J Neurosci. 2015 Jun 24;35(25):9369-80. doi: 10.1523/JNEUROSCI.5054-14.2015. J Neurosci. 2015. PMID: 26109660 Free PMC article.
References
-
- Liberman MC, Dodds LW. Single-neuron labeling and chronic cochlear pathology. III. Stereocilia damage and alterations of threshold tuning curves. Hear Res. 1984;16(1):55–74. - PubMed
-
- Liberman MC, Dodds LW. Single-neuron labeling and chronic cochlear pathology. II. Stereocilia damage and alterations of spontaneous discharge rates. Hear Res. 1984;16(1):43–53. - PubMed
-
- Shepherd RK, Roberts LA, Paolini AG. Long-term sensorineural hearing loss induces functional changes in the rat auditory nerve. Eur J Neurosci. 2004;20:3131–3140. - PubMed
-
- Pujol R, Puel J-L. Excitotoxicity, synaptic repair, and functional recovery in the mammalian cochlea: A review of recent findings. Ann N Y Acad Sci. 1999;884:249–254. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
