

Dasatinib as a bone-modifying agent: anabolic and anti-resorptive effects
- PMID: 22539950
- PMCID: PMC3335111
- DOI: 10.1371/journal.pone.0034914
Dasatinib as a bone-modifying agent: anabolic and anti-resorptive effects
Abstract
Background: Bone loss, in malignant or non-malignant diseases, is caused by increased osteoclast resorption and/or reduced osteoblast bone formation, and is commonly associated with skeletal complications. Thus, there is a need to identify new agents capable of influencing bone remodeling. We aimed to further pre-clinically evaluate the effects of dasatinib (BMS-354825), a multitargeted tyrosine kinase inhibitor, on osteoblast and osteoclast differentiation and function.
Methods: For studies on osteoblasts, primary human bone marrow mensenchymal stem cells (hMSCs) together with the hMSC-TERT and the MG-63 cell lines were employed. Osteoclasts were generated from peripheral blood mononuclear cells (PBMC) of healthy volunteers. Skeletally-immature CD1 mice were used in the in vivo model.
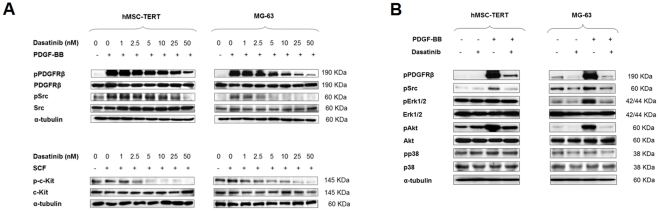
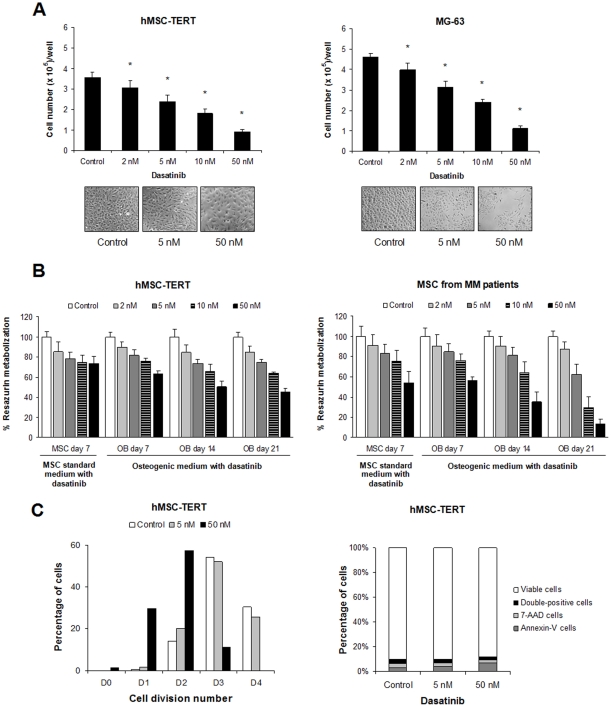
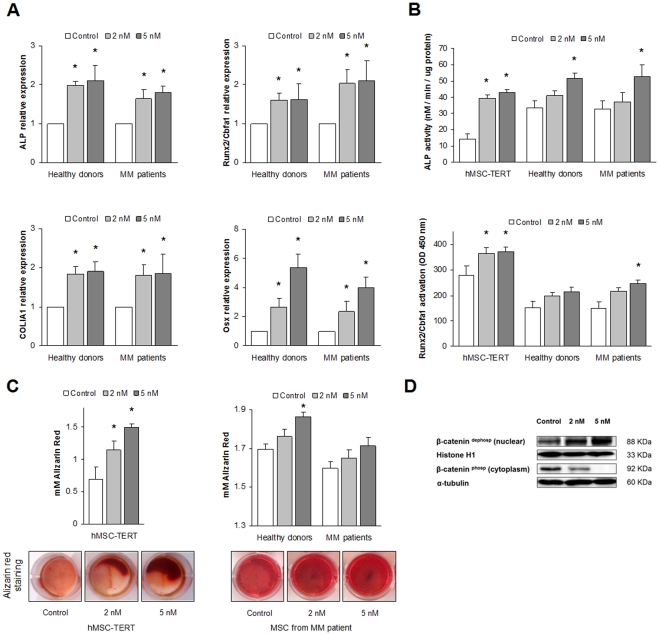
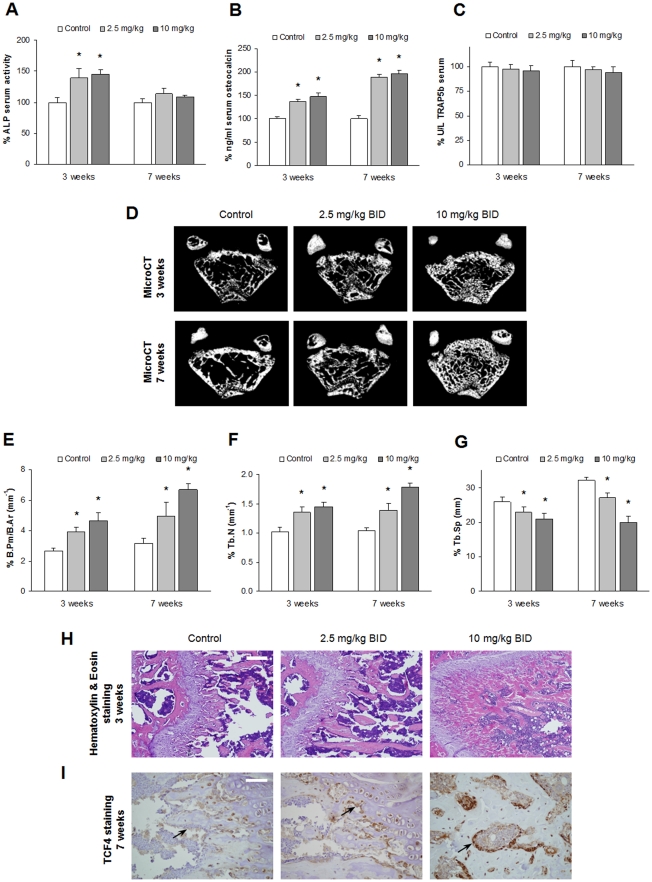
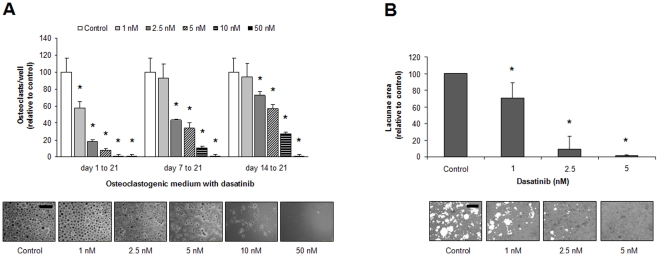
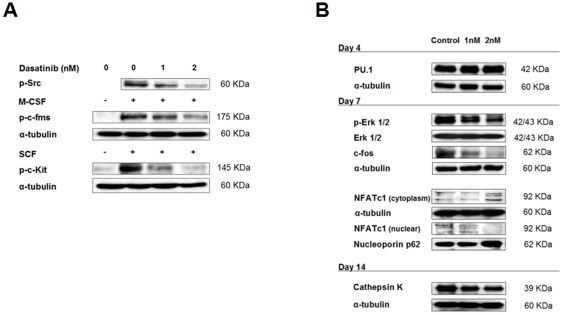
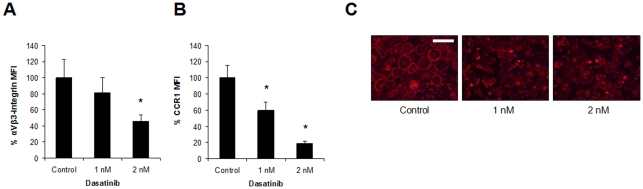
Results: Dasatinib inhibited the platelet derived growth factor receptor-β (PDGFR-β), c-Src and c-Kit phosphorylation in hMSC-TERT and MG-63 cell lines, which was associated with decreased cell proliferation and activation of canonical Wnt signaling. Treatment of MSCs from healthy donors, but also from multiple myeloma patients with low doses of dasatinib (2-5 nM), promoted its osteogenic differentiation and matrix mineralization. The bone anabolic effect of dasatinib was also observed in vivo by targeting endogenous osteoprogenitors, as assessed by elevated serum levels of bone formation markers, and increased trabecular microarchitecture and number of osteoblast-like cells. By in vitro exposure of hemopoietic progenitors to a similar range of dasatinib concentrations (1-2 nM), novel biological sequelae relative to inhibition of osteoclast formation and resorptive function were identified, including F-actin ring disruption, reduced levels of c-Fos and of nuclear factor of activated T cells 1 (NFATc1) in the nucleus, together with lowered cathepsin K, αVβ3 integrin and CCR1 expression.
Conclusions: Low dasatinib concentrations show convergent bone anabolic and reduced bone resorption effects, which suggests its potential use for the treatment of bone diseases such as osteoporosis, osteolytic bone metastasis and myeloma bone disease.
Conflict of interest statement
Figures
Similar articles
-
The tyrosine kinase inhibitor dasatinib dysregulates bone remodeling through inhibition of osteoclasts in vivo.J Bone Miner Res. 2010 Aug;25(8):1759-70. doi: 10.1002/jbmr.85. J Bone Miner Res. 2010. PMID: 20225261
-
Dasatinib inhibits both osteoclast activation and prostate cancer PC-3-cell-induced osteoclast formation.Cancer Biol Ther. 2009 Nov;8(22):2153-9. doi: 10.4161/cbt.8.22.9770. Epub 2009 Nov 8. Cancer Biol Ther. 2009. PMID: 19855158 Free PMC article.
-
WHI-131 Promotes Osteoblast Differentiation and Prevents Osteoclast Formation and Resorption in Mice.J Bone Miner Res. 2016 Feb;31(2):403-15. doi: 10.1002/jbmr.2612. Epub 2015 Aug 29. J Bone Miner Res. 2016. PMID: 26255791
-
Biological aspects of altered bone remodeling in multiple myeloma and possibilities of pharmacological intervention.Dan Med Bull. 2011 May;58(5):B4277. Dan Med Bull. 2011. PMID: 21535989 Review.
-
The osteoclast and its unique cytoskeleton.Ann N Y Acad Sci. 2011 Dec;1240:14-7. doi: 10.1111/j.1749-6632.2011.06283.x. Ann N Y Acad Sci. 2011. PMID: 22172034 Review.
Cited by
-
Age-associated callus senescent cells produce TGF-β1 that inhibits fracture healing in aged mice.J Clin Invest. 2022 Apr 15;132(8):e148073. doi: 10.1172/JCI148073. J Clin Invest. 2022. PMID: 35426372 Free PMC article.
-
Treatment Effects of the Second-Generation Tyrosine Kinase Inhibitor Dasatinib on Autoimmune Arthritis.Front Immunol. 2019 Jan 10;9:3133. doi: 10.3389/fimmu.2018.03133. eCollection 2018. Front Immunol. 2019. PMID: 30687331 Free PMC article.
-
Dasatinib prevents skeletal metastasis of osteotropic MDA-MB-231 cells in a xenograft mouse model.Arch Gynecol Obstet. 2020 Jun;301(6):1493-1502. doi: 10.1007/s00404-020-05496-4. Epub 2020 Mar 14. Arch Gynecol Obstet. 2020. PMID: 32170411
-
Transcriptomic profile induced in bone marrow mesenchymal stromal cells after interaction with multiple myeloma cells: implications in myeloma progression and myeloma bone disease.Oncotarget. 2014 Sep 30;5(18):8284-305. doi: 10.18632/oncotarget.2058. Oncotarget. 2014. PMID: 25268740 Free PMC article.
-
Receptor tyrosine kinases: Characterisation, mechanism of action and therapeutic interests for bone cancers.J Bone Oncol. 2015 Jan 23;4(1):1-12. doi: 10.1016/j.jbo.2015.01.001. eCollection 2015 Mar. J Bone Oncol. 2015. PMID: 26579483 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
