

DNA damage-inducible SUMOylation of HERC2 promotes RNF8 binding via a novel SUMO-binding Zinc finger
- PMID: 22508508
- PMCID: PMC3328386
- DOI: 10.1083/jcb.201106152
DNA damage-inducible SUMOylation of HERC2 promotes RNF8 binding via a novel SUMO-binding Zinc finger
Abstract
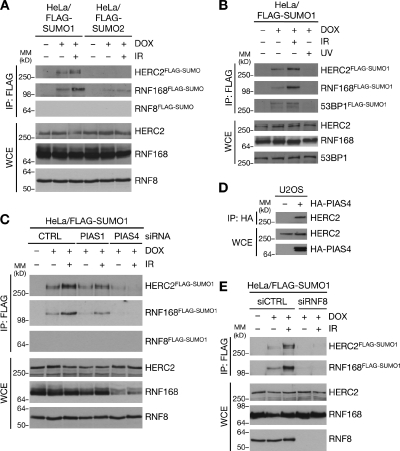
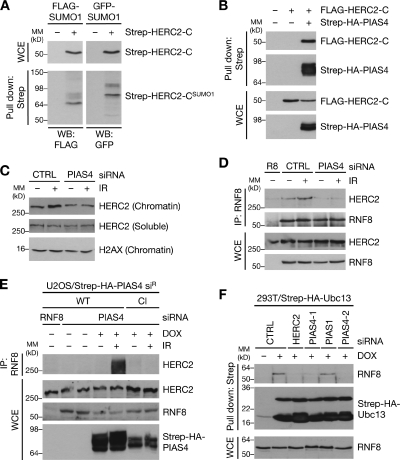
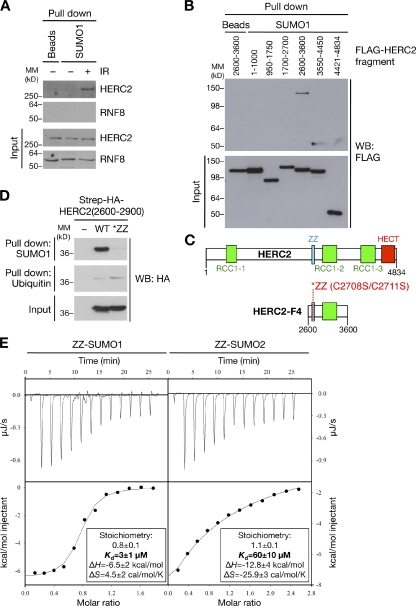
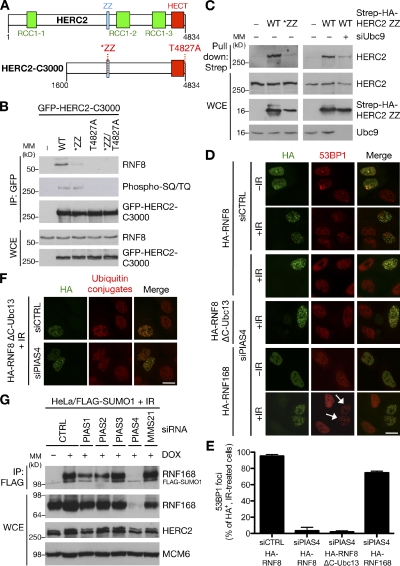
Nonproteolytic ubiquitylation of chromatin surrounding deoxyribonucleic acid (DNA) double-strand breaks (DSBs) by the RNF8/RNF168/HERC2 ubiquitin ligases facilitates restoration of genome integrity by licensing chromatin to concentrate genome caretaker proteins near the lesions. In parallel, SUMOylation of so-far elusive upstream DSB regulators is also required for execution of this ubiquitin-dependent chromatin response. We show that HERC2 and RNF168 are novel DNA damage-dependent SUMOylation targets in human cells. In response to DSBs, both HERC2 and RNF168 were specifically modified with SUMO1 at DSB sites in a manner dependent on the SUMO E3 ligase PIAS4. SUMOylation of HERC2 was required for its DSB-induced association with RNF8 and for stabilizing the RNF8-Ubc13 complex. We also demonstrate that the ZZ Zinc finger in HERC2 defined a novel SUMO-specific binding module, which together with its concomitant SUMOylation and T4827 phosphorylation promoted binding to RNF8. Our findings provide novel insight into the regulatory complexity of how ubiquitylation and SUMOylation cooperate to orchestrate protein interactions with DSB repair foci.
Figures
Similar articles
-
The ubiquitin- and SUMO-dependent signaling response to DNA double-strand breaks.FEBS Lett. 2011 Sep 16;585(18):2914-9. doi: 10.1016/j.febslet.2011.05.056. Epub 2011 Jun 12. FEBS Lett. 2011. PMID: 21664912 Review.
-
Mammalian SUMO E3-ligases PIAS1 and PIAS4 promote responses to DNA double-strand breaks.Nature. 2009 Dec 17;462(7275):935-9. doi: 10.1038/nature08657. Nature. 2009. PMID: 20016603 Free PMC article.
-
Histone H1 couples initiation and amplification of ubiquitin signalling after DNA damage.Nature. 2015 Nov 19;527(7578):389-93. doi: 10.1038/nature15401. Epub 2015 Oct 21. Nature. 2015. PMID: 26503038
-
Human RNF169 is a negative regulator of the ubiquitin-dependent response to DNA double-strand breaks.J Cell Biol. 2012 Apr 16;197(2):189-99. doi: 10.1083/jcb.201109100. Epub 2012 Apr 9. J Cell Biol. 2012. PMID: 22492721 Free PMC article.
-
Regulatory ubiquitylation in response to DNA double-strand breaks.DNA Repair (Amst). 2009 Apr 5;8(4):436-43. doi: 10.1016/j.dnarep.2009.01.013. Epub 2009 Feb 18. DNA Repair (Amst). 2009. PMID: 19230794 Review.
Cited by
-
SUMO and ubiquitin-dependent XPC exchange drives nucleotide excision repair.Nat Commun. 2015 Jul 7;6:7499. doi: 10.1038/ncomms8499. Nat Commun. 2015. PMID: 26151477 Free PMC article.
-
Chromatin Ubiquitination Guides DNA Double Strand Break Signaling and Repair.Front Cell Dev Biol. 2022 Jul 5;10:928113. doi: 10.3389/fcell.2022.928113. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35865631 Free PMC article. Review.
-
HERC2-USP20 axis regulates DNA damage checkpoint through Claspin.Nucleic Acids Res. 2014 Dec 1;42(21):13110-21. doi: 10.1093/nar/gku1034. Epub 2014 Oct 29. Nucleic Acids Res. 2014. PMID: 25355518 Free PMC article.
-
Spatiotemporal regulation of posttranslational modifications in the DNA damage response.EMBO J. 2016 Jan 4;35(1):6-23. doi: 10.15252/embj.201592595. Epub 2015 Dec 1. EMBO J. 2016. PMID: 26628622 Free PMC article. Review.
-
Role of the CBP catalytic core in intramolecular SUMOylation and control of histone H3 acetylation.Proc Natl Acad Sci U S A. 2017 Jul 3;114(27):E5335-E5342. doi: 10.1073/pnas.1703105114. Epub 2017 Jun 19. Proc Natl Acad Sci U S A. 2017. PMID: 28630323 Free PMC article.
References
-
- Bekker-Jensen S., Rendtlew Danielsen J., Fugger K., Gromova I., Nerstedt A., Lukas C., Bartek J., Lukas J., Mailand N. 2010. HERC2 coordinates ubiquitin-dependent assembly of DNA repair factors on damaged chromosomes. Nat. Cell Biol. 12:80–86: 1–12 (published erratum appears in Nat. Cell Biol. 2010. 12:412) 10.1038/ncb2008 - DOI - PubMed
-
- Devgan S.S., Sanal O., Doil C., Nakamura K., Nahas S.A., Pettijohn K., Bartek J., Lukas C., Lukas J., Gatti R.A. 2011. Homozygous deficiency of ubiquitin-ligase ring-finger protein RNF168 mimics the radiosensitivity syndrome of ataxia-telangiectasia. Cell Death Differ. 18:1500–1506 10.1038/cdd.2011.18 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
