

Coordinated regulation of accessory genetic elements produces cyclic di-nucleotides for V. cholerae virulence
- PMID: 22500802
- PMCID: PMC3620040
- DOI: 10.1016/j.cell.2012.01.053
Coordinated regulation of accessory genetic elements produces cyclic di-nucleotides for V. cholerae virulence
Abstract
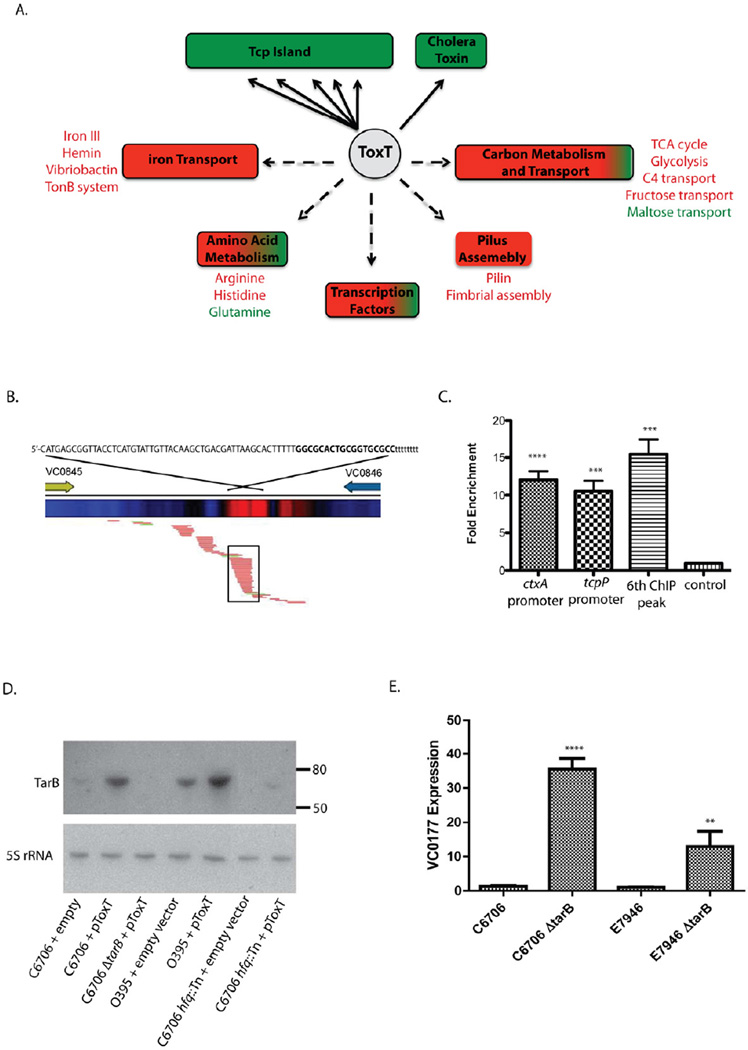
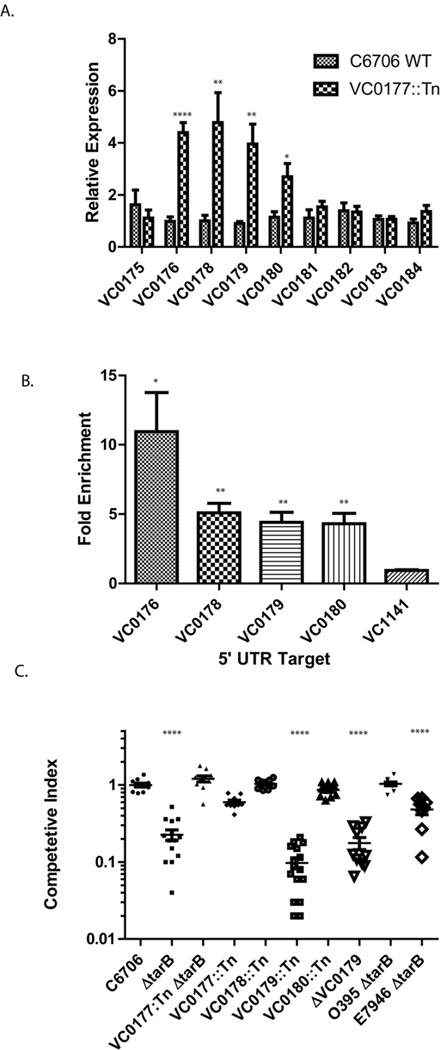
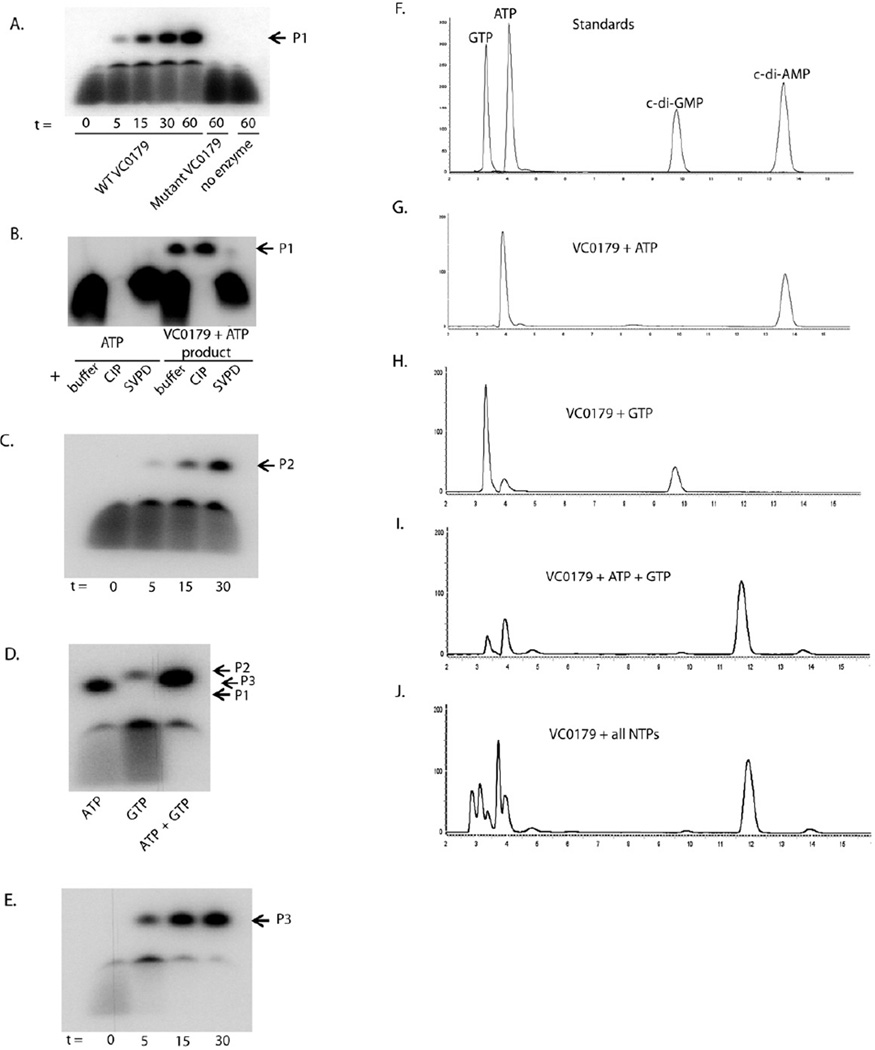
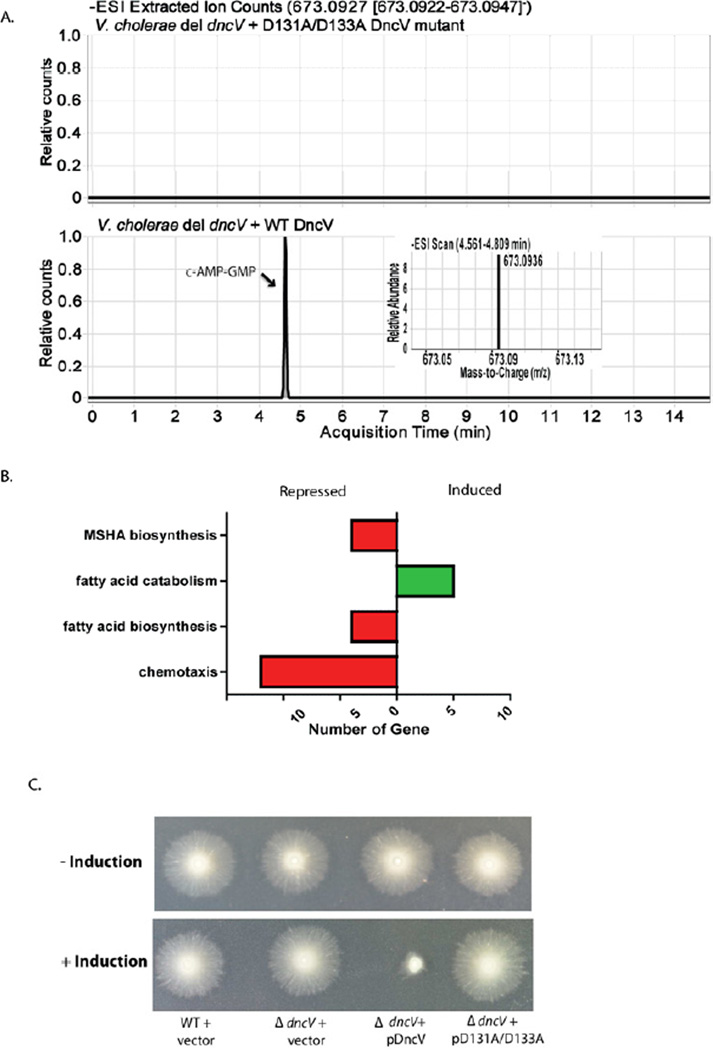
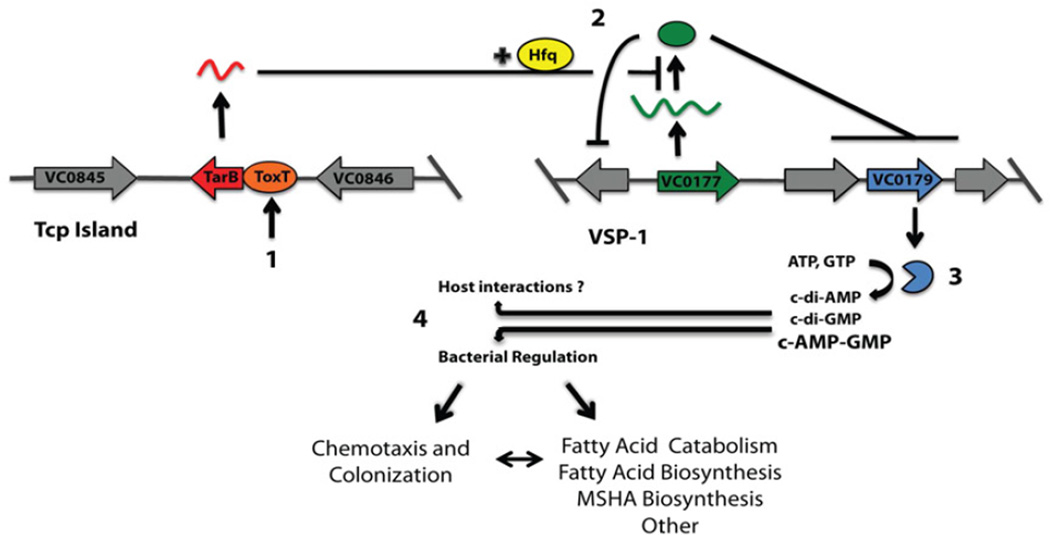
The function of the Vibrio 7(th) pandemic island-1 (VSP-1) in cholera pathogenesis has remained obscure. Utilizing chromatin immunoprecipitation sequencing and RNA sequencing to map the regulon of the master virulence regulator ToxT, we identify a TCP island-encoded small RNA that reduces the expression of a previously unrecognized VSP-1-encoded transcription factor termed VspR. VspR modulates the expression of several VSP-1 genes including one that encodes a novel class of di-nucleotide cyclase (DncV), which preferentially synthesizes a previously undescribed hybrid cyclic AMP-GMP molecule. We show that DncV is required for efficient intestinal colonization and downregulates V. cholerae chemotaxis, a phenotype previously associated with hyperinfectivity. This pathway couples the actions of previously disparate genomic islands, defines VSP-1 as a pathogenicity island in V. cholerae, and implicates its occurrence in 7(th) pandemic strains as a benefit for host adaptation through the production of a regulatory cyclic di-nucleotide.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Bacterial virulence: A new pathogenicity island for Vibrio.Nat Rev Microbiol. 2012 May 8;10(6):376-7. doi: 10.1038/nrmicro2799. Nat Rev Microbiol. 2012. PMID: 22565129 No abstract available.
Similar articles
-
Vibrio cholerae ToxR downregulates virulence factor production in response to cyclo(Phe-Pro).mBio. 2013 Aug 27;4(5):e00366-13. doi: 10.1128/mBio.00366-13. mBio. 2013. PMID: 23982069 Free PMC article.
-
The Vibrio cholerae virulence regulatory cascade controls glucose uptake through activation of TarA, a small regulatory RNA.Mol Microbiol. 2010 Dec;78(5):1171-81. doi: 10.1111/j.1365-2958.2010.07397.x. Epub 2010 Sep 30. Mol Microbiol. 2010. PMID: 21091503 Free PMC article.
-
Distribution of genes for virulence and ecological fitness among diverse Vibrio cholerae population in a cholera endemic area: tracking the evolution of pathogenic strains.DNA Cell Biol. 2008 Jul;27(7):347-55. doi: 10.1089/dna.2008.0737. DNA Cell Biol. 2008. PMID: 18462070 Free PMC article.
-
Multiple intraintestinal signals coordinate the regulation of Vibrio cholerae virulence determinants.Pathog Dis. 2018 Feb 1;76(1). doi: 10.1093/femspd/ftx126. Pathog Dis. 2018. PMID: 29315383 Review.
-
Regulation of virulence in Vibrio cholerae: the ToxR regulon.Future Microbiol. 2007 Jun;2(3):335-44. doi: 10.2217/17460913.2.3.335. Future Microbiol. 2007. PMID: 17661707 Review.
Cited by
-
Bacteria sting viral invaders.Nature. 2020 Oct;586(7829):363-364. doi: 10.1038/d41586-020-02712-8. Nature. 2020. PMID: 32989308 No abstract available.
-
Ubiquitin-like conjugation by bacterial cGAS enhances anti-phage defence.Nature. 2023 Apr;616(7956):326-331. doi: 10.1038/s41586-023-05862-7. Epub 2023 Feb 27. Nature. 2023. PMID: 36848932 Free PMC article.
-
Recombinant BCG With Bacterial Signaling Molecule Cyclic di-AMP as Endogenous Adjuvant Induces Elevated Immune Responses After Mycobacterium tuberculosis Infection.Front Immunol. 2019 Jul 3;10:1519. doi: 10.3389/fimmu.2019.01519. eCollection 2019. Front Immunol. 2019. PMID: 31333655 Free PMC article.
-
The Innate Antiviral Response in Animals: An Evolutionary Perspective from Flagellates to Humans.Viruses. 2019 Aug 16;11(8):758. doi: 10.3390/v11080758. Viruses. 2019. PMID: 31426357 Free PMC article. Review.
-
Essentiality of the Maltase AmlE in Maltose Utilization and Its Transcriptional Regulation by the Repressor AmlR in the Acarbose-Producing Bacterium Actinoplanes sp. SE50/110.Front Microbiol. 2019 Oct 29;10:2448. doi: 10.3389/fmicb.2019.02448. eCollection 2019. Front Microbiol. 2019. PMID: 31736895 Free PMC article.
References
-
- Brennan RG, Link TM. Hfq structure, function and ligand binding. Curr Opin Microbiol. 2007;10:125–133. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
