

An inverse relationship to germline transcription defines centromeric chromatin in C. elegans
- PMID: 22495302
- PMCID: PMC3538161
- DOI: 10.1038/nature10973
An inverse relationship to germline transcription defines centromeric chromatin in C. elegans
Abstract
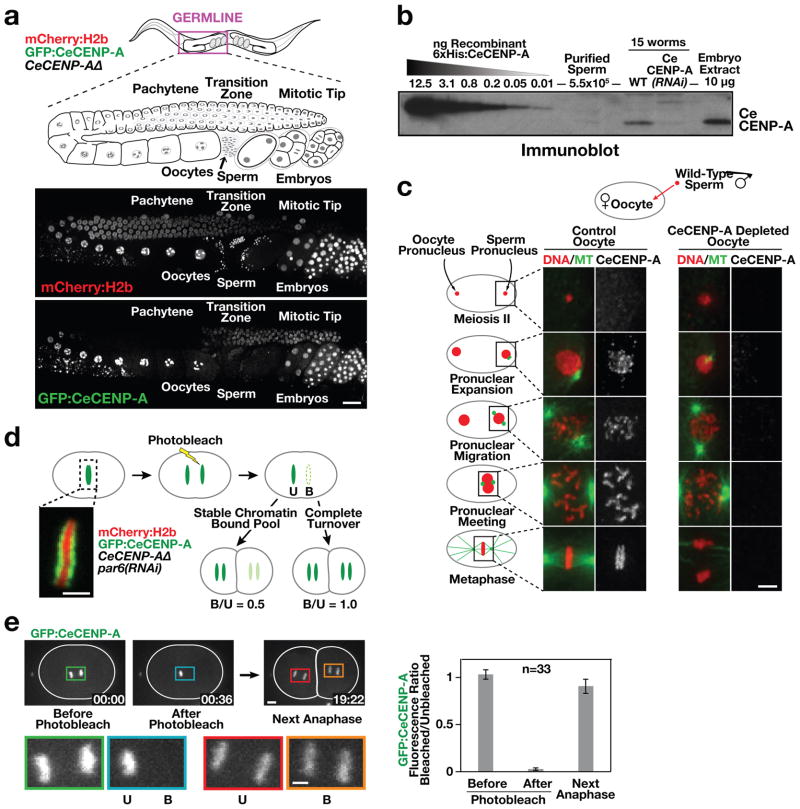
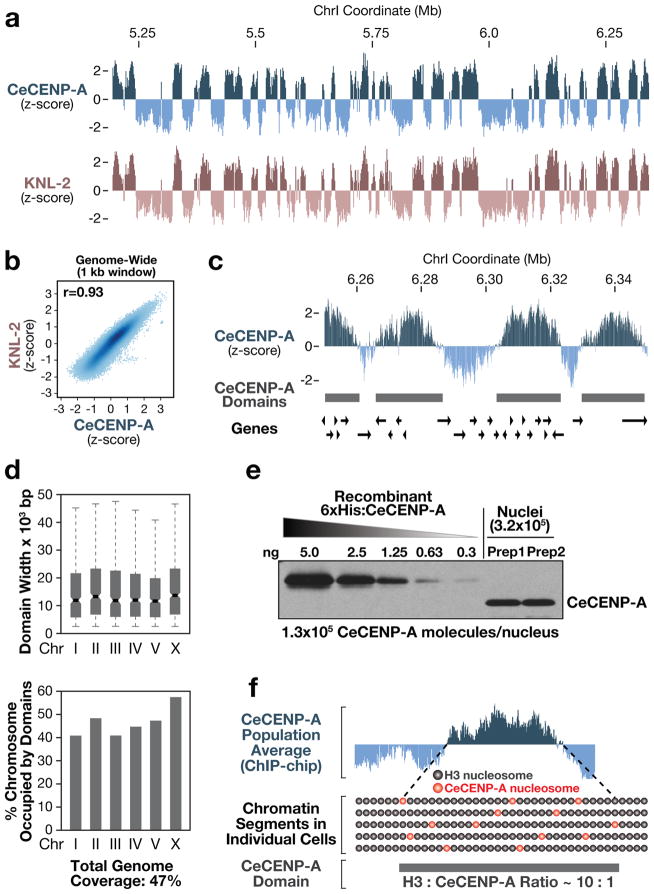
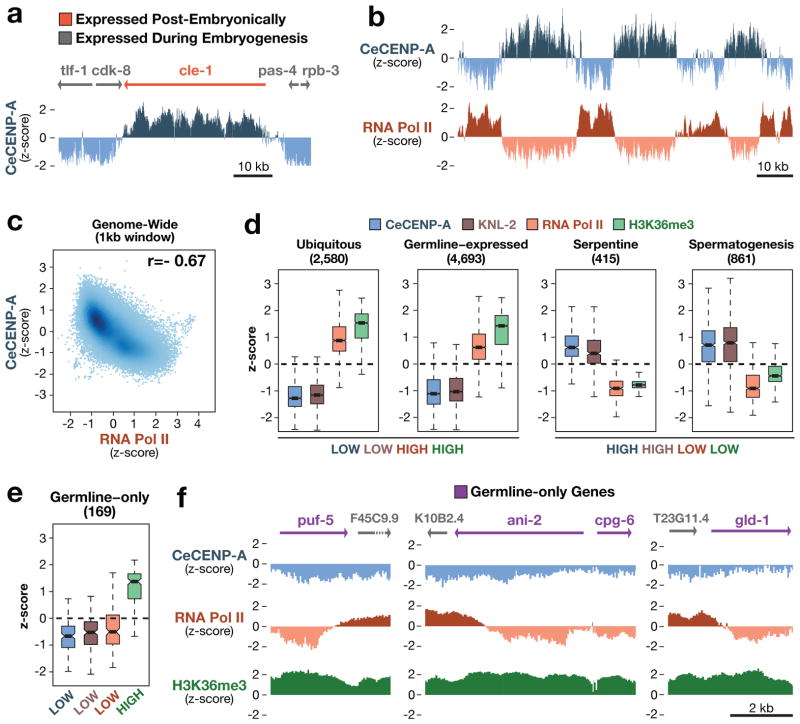
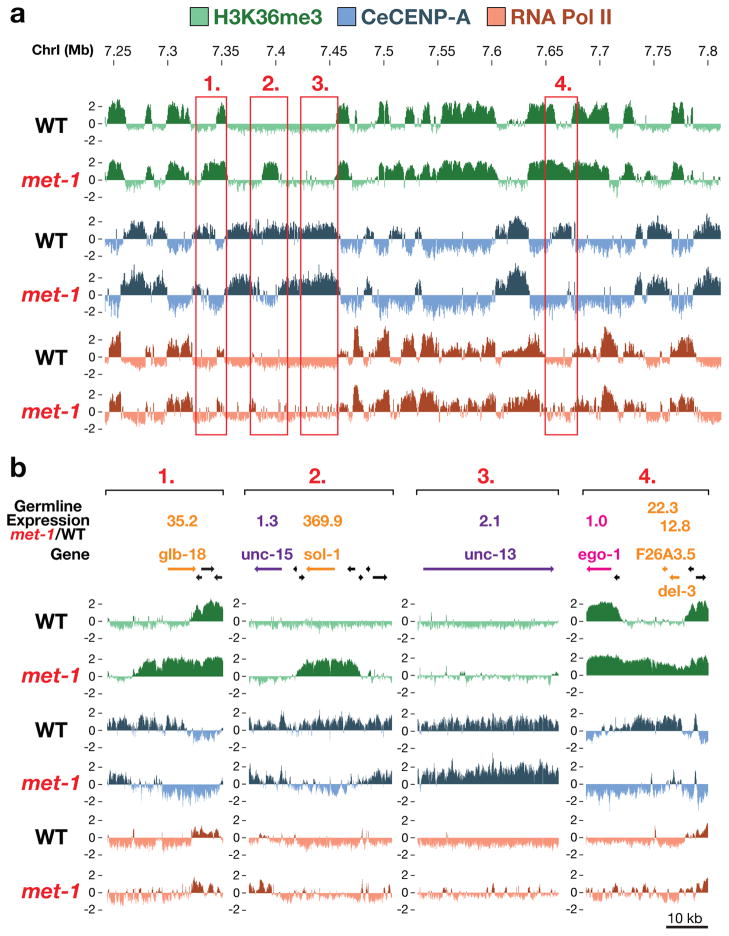
Centromeres are chromosomal loci that direct segregation of the genome during cell division. The histone H3 variant CENP-A (also known as CenH3) defines centromeres in monocentric organisms, which confine centromere activity to a discrete chromosomal region, and holocentric organisms, which distribute centromere activity along the chromosome length. Because the highly repetitive DNA found at most centromeres is neither necessary nor sufficient for centromere function, stable inheritance of CENP-A nucleosomal chromatin is postulated to propagate centromere identity epigenetically. Here, we show that in the holocentric nematode Caenorhabditis elegans pre-existing CENP-A nucleosomes are not necessary to guide recruitment of new CENP-A nucleosomes. This is indicated by lack of CENP-A transmission by sperm during fertilization and by removal and subsequent reloading of CENP-A during oogenic meiotic prophase. Genome-wide mapping of CENP-A location in embryos and quantification of CENP-A molecules in nuclei revealed that CENP-A is incorporated at low density in domains that cumulatively encompass half the genome. Embryonic CENP-A domains are established in a pattern inverse to regions that are transcribed in the germline and early embryo, and ectopic transcription of genes in a mutant germline altered the pattern of CENP-A incorporation in embryos. Furthermore, regions transcribed in the germline but not embryos fail to incorporate CENP-A throughout embryogenesis. We propose that germline transcription defines genomic regions that exclude CENP-A incorporation in progeny, and that zygotic transcription during early embryogenesis remodels and reinforces this basal pattern. These findings link centromere identity to transcription and shed light on the evolutionary plasticity of centromeres.
Figures
Similar articles
-
Transgenerational inheritance of centromere identity requires the CENP-A N-terminal tail in the C. elegans maternal germ line.PLoS Biol. 2021 Jul 6;19(7):e3000968. doi: 10.1371/journal.pbio.3000968. eCollection 2021 Jul. PLoS Biol. 2021. PMID: 34228701 Free PMC article.
-
RbAp46/48(LIN-53) Is Required for Holocentromere Assembly in Caenorhabditis elegans.Cell Rep. 2016 Mar 1;14(8):1819-28. doi: 10.1016/j.celrep.2016.01.065. Epub 2016 Feb 18. Cell Rep. 2016. PMID: 26904949
-
Functional genomics identifies a Myb domain-containing protein family required for assembly of CENP-A chromatin.J Cell Biol. 2007 Mar 12;176(6):757-63. doi: 10.1083/jcb.200701065. Epub 2007 Mar 5. J Cell Biol. 2007. PMID: 17339379 Free PMC article.
-
Maternal inheritance of centromeres through the germline.Curr Top Dev Biol. 2020;140:35-54. doi: 10.1016/bs.ctdb.2020.03.004. Epub 2020 Apr 25. Curr Top Dev Biol. 2020. PMID: 32591081 Review.
-
The ins and outs of CENP-A: Chromatin dynamics of the centromere-specific histone.Semin Cell Dev Biol. 2023 Feb 15;135:24-34. doi: 10.1016/j.semcdb.2022.04.003. Epub 2022 Apr 11. Semin Cell Dev Biol. 2023. PMID: 35422390 Review.
Cited by
-
Comparative analysis of tandem repeats from hundreds of species reveals unique insights into centromere evolution.Genome Biol. 2013 Jan 30;14(1):R10. doi: 10.1186/gb-2013-14-1-r10. Genome Biol. 2013. PMID: 23363705 Free PMC article.
-
Phosphorylation of RNA polymerase II is independent of P-TEFb in the C. elegans germline.Development. 2013 Sep;140(17):3703-13. doi: 10.1242/dev.095778. Epub 2013 Jul 31. Development. 2013. PMID: 23903194 Free PMC article.
-
A multitasking Argonaute: exploring the many facets of C. elegans CSR-1.Chromosome Res. 2013 Dec;21(6-7):573-86. doi: 10.1007/s10577-013-9383-7. Chromosome Res. 2013. PMID: 24178449 Review.
-
Genome-wide analysis links emerin to neuromuscular junction activity in Caenorhabditis elegans.Genome Biol. 2014 Feb 3;15(2):R21. doi: 10.1186/gb-2014-15-2-r21. Genome Biol. 2014. PMID: 24490688 Free PMC article.
-
Comparative analysis of metazoan chromatin organization.Nature. 2014 Aug 28;512(7515):449-52. doi: 10.1038/nature13415. Nature. 2014. PMID: 25164756 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
