

Cell Membrane-associated heparan sulfate is a receptor for prototype foamy virus in human, monkey, and rodent cells
- PMID: 22434139
- PMCID: PMC3369305
- DOI: 10.1038/mt.2012.41
Cell Membrane-associated heparan sulfate is a receptor for prototype foamy virus in human, monkey, and rodent cells
Abstract
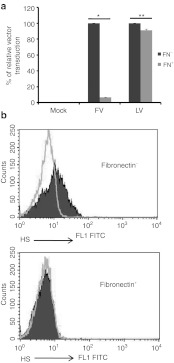
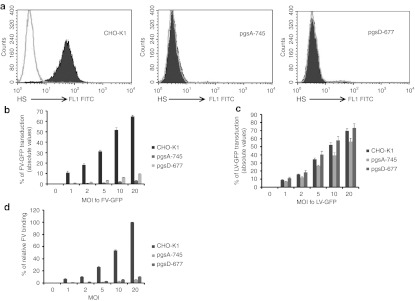
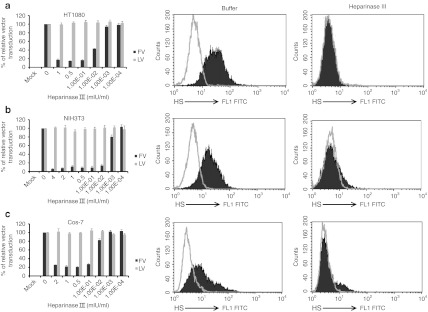
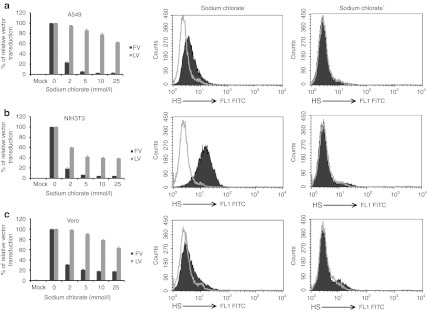
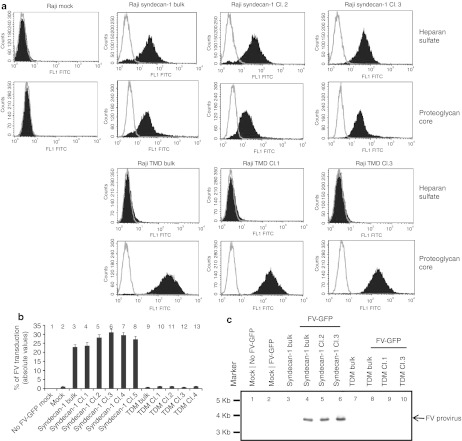
Foamy viruses (FVs) (spumaretroviruses) are good alternative to retroviruses as gene therapy vector. Despite four decades since the discovery of FV, its receptor molecule is still unknown. FV vector transduction of human CD34(+) cells was inhibited by culture with fibronectin. Because fibronectin contains heparin-binding domain, the interactions of fibronectin with heparan sulfate (HS) on cells might be inhibitory to FV transduction. These observations led us to investigate whether HS is a receptor for FV. Two mutant CHO cell lines (but not parental wild type) lacking cell surface HS but not chondroitin sulfate (CS) were largely resistant to FV attachment and transduction. Inhibition of HS expression using enzymes or chemicals greatly reduced FV transduction in human, monkey, and rodent cells. Raji cells, which lack HS and were largely resistant to FV, were rendered more permissive through ectopic expression of syndecan-1, which contains HS. In contrast, mutant syndecan-1-expressing cells were largely resistant to FV. Our findings indicate that cellular HS is a receptor for FV. Identifying FV receptor will enable better understanding of its entry process and optimal use as gene therapy vector to treat inherited and pathogenic diseases.
Figures
Comment in
-
Advances in foamy virus vector technology and disease correction could speed the path to clinical application.Mol Ther. 2012 Jun;20(6):1105-7. doi: 10.1038/mt.2012.97. Mol Ther. 2012. PMID: 22652999 Free PMC article. No abstract available.
Similar articles
-
Heparan sulfate is an attachment factor for foamy virus entry.J Virol. 2012 Sep;86(18):10028-35. doi: 10.1128/JVI.00051-12. Epub 2012 Jul 11. J Virol. 2012. PMID: 22787203 Free PMC article.
-
Adaptation of Sindbis virus to BHK cells selects for use of heparan sulfate as an attachment receptor.J Virol. 1998 Sep;72(9):7357-66. doi: 10.1128/JVI.72.9.7357-7366.1998. J Virol. 1998. PMID: 9696832 Free PMC article.
-
The furin protease cleavage recognition sequence of Sindbis virus PE2 can mediate virion attachment to cell surface heparan sulfate.J Virol. 1999 Aug;73(8):6299-306. doi: 10.1128/JVI.73.8.6299-6306.1999. J Virol. 1999. PMID: 10400721 Free PMC article.
-
Role of heparan sulfate for attachment and entry of tick-borne encephalitis virus.Virology. 2003 Mar 30;308(1):92-100. doi: 10.1016/s0042-6822(02)00097-1. Virology. 2003. PMID: 12706093
-
Close and focal contact adhesions of fibroblasts to a fibronectin-containing matrix.Fed Proc. 1985 Feb;44(2):394-403. Fed Proc. 1985. PMID: 3917945 Review.
Cited by
-
Early events in foamy virus-host interaction and intracellular trafficking.Viruses. 2013 Apr 8;5(4):1055-74. doi: 10.3390/v5041055. Viruses. 2013. PMID: 23567621 Free PMC article. Review.
-
Heparanase Blockade as a Novel Dual-Targeting Therapy for COVID-19.J Virol. 2022 Apr 13;96(7):e0005722. doi: 10.1128/jvi.00057-22. Epub 2022 Mar 23. J Virol. 2022. PMID: 35319225 Free PMC article.
-
Efficient production of inhibitor-free foamy virus glycoprotein-containing retroviral vectors by proteoglycan-deficient packaging cells.Mol Ther Methods Clin Dev. 2022 Aug 1;26:394-412. doi: 10.1016/j.omtm.2022.07.004. eCollection 2022 Sep 8. Mol Ther Methods Clin Dev. 2022. PMID: 36034773 Free PMC article.
-
Foamy Viruses, Bet, and APOBEC3 Restriction.Viruses. 2021 Mar 18;13(3):504. doi: 10.3390/v13030504. Viruses. 2021. PMID: 33803830 Free PMC article. Review.
-
IL-2 Inducible Kinase ITK is Critical for HIV-1 Infection of Jurkat T-cells.Sci Rep. 2018 Feb 16;8(1):3217. doi: 10.1038/s41598-018-21344-7. Sci Rep. 2018. PMID: 29453458 Free PMC article.
References
-
- Linial M.2007Foamy viruses Fields BN, Knipe DM., and, Howley PM.eds). Fields Virology5th edn, vol. 1Lippincott Williams & Wilkins: Philadelphia; 2245–2262.
-
- Rethwilm A. Foamy virus vectors: an awaited alternative to gammaretro- and lentiviral vectors. Curr Gene Ther. 2007;7:261–271. - PubMed
-
- Rethwilm A. Molecular biology of foamy viruses. Med Microbiol Immunol. 2010;199:197–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
