

Tumor development is associated with decrease of TET gene expression and 5-methylcytosine hydroxylation
- PMID: 22391558
- PMCID: PMC3897214
- DOI: 10.1038/onc.2012.67
Tumor development is associated with decrease of TET gene expression and 5-methylcytosine hydroxylation
Abstract
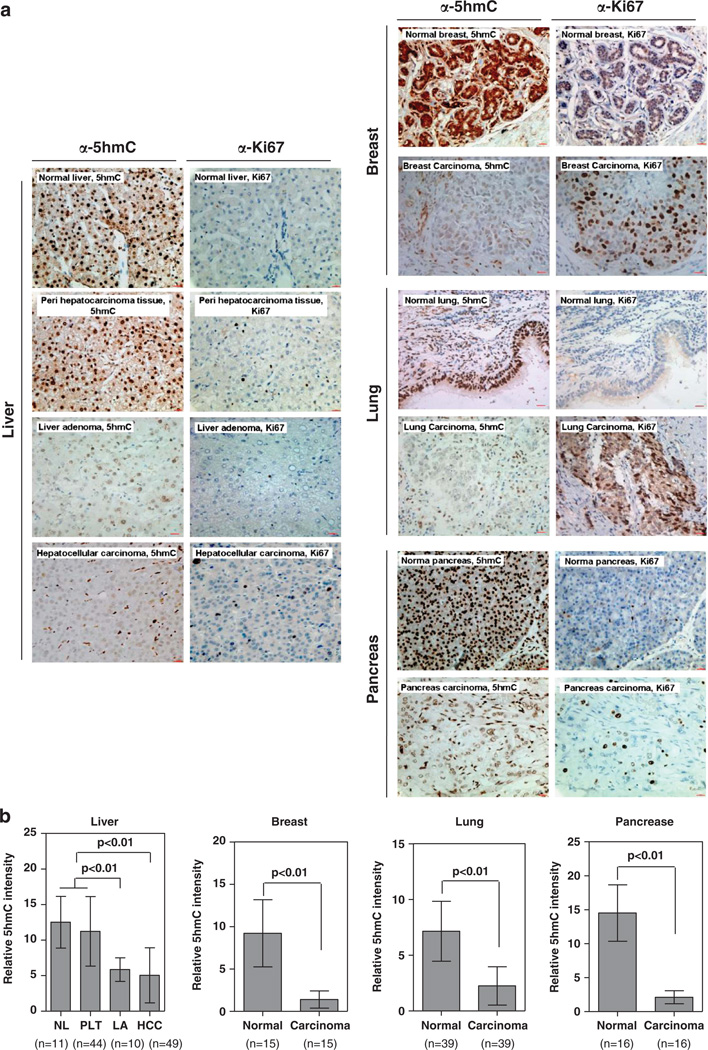
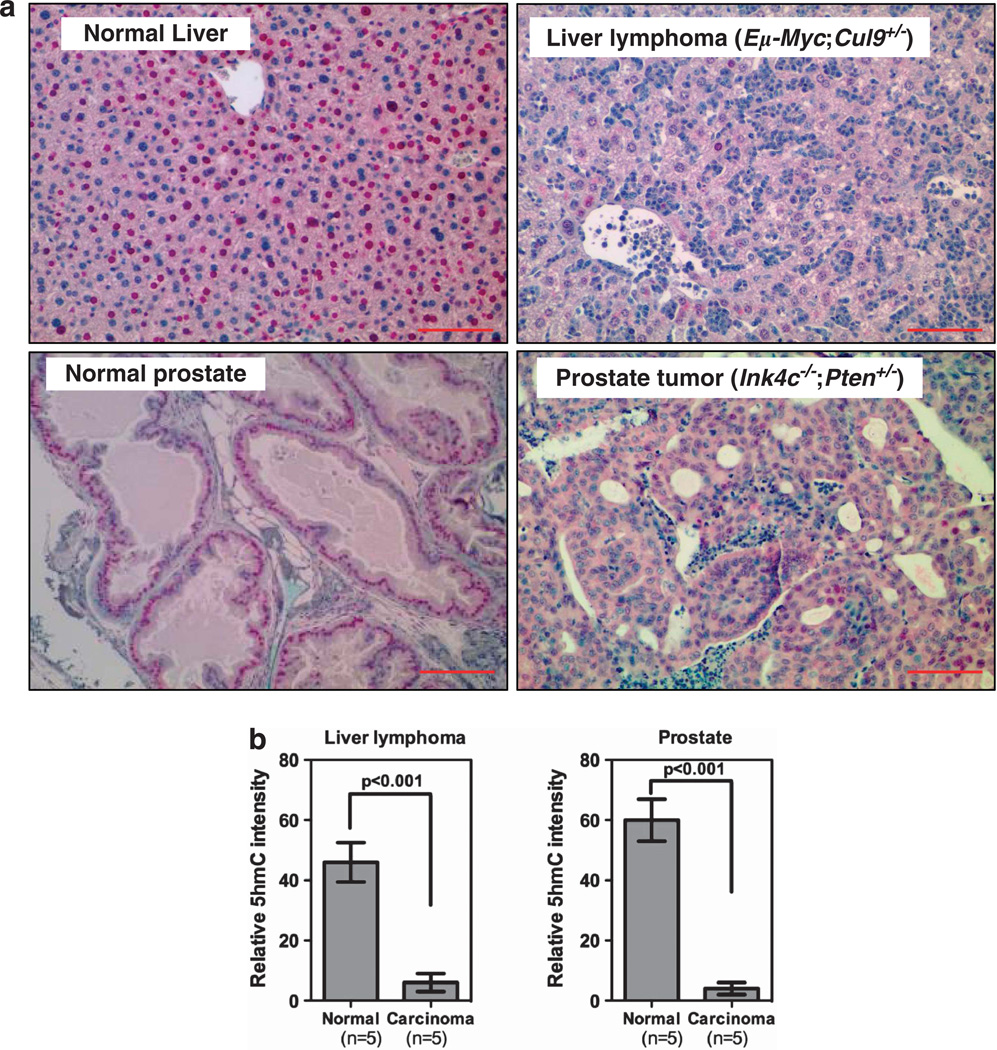
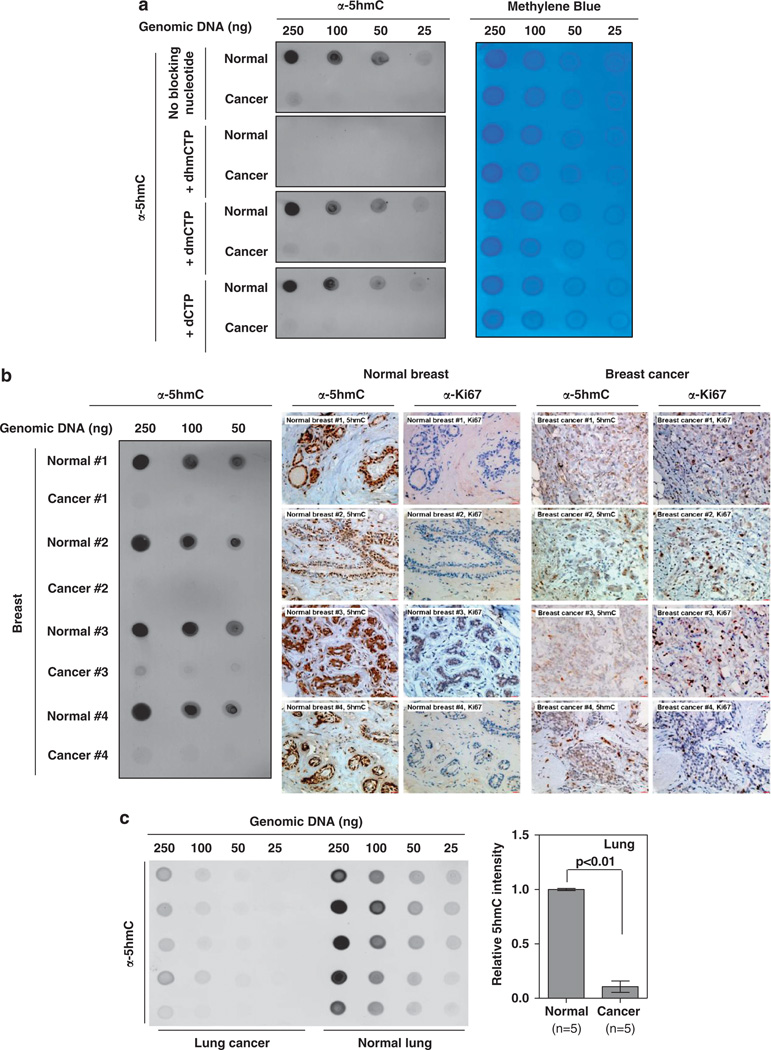
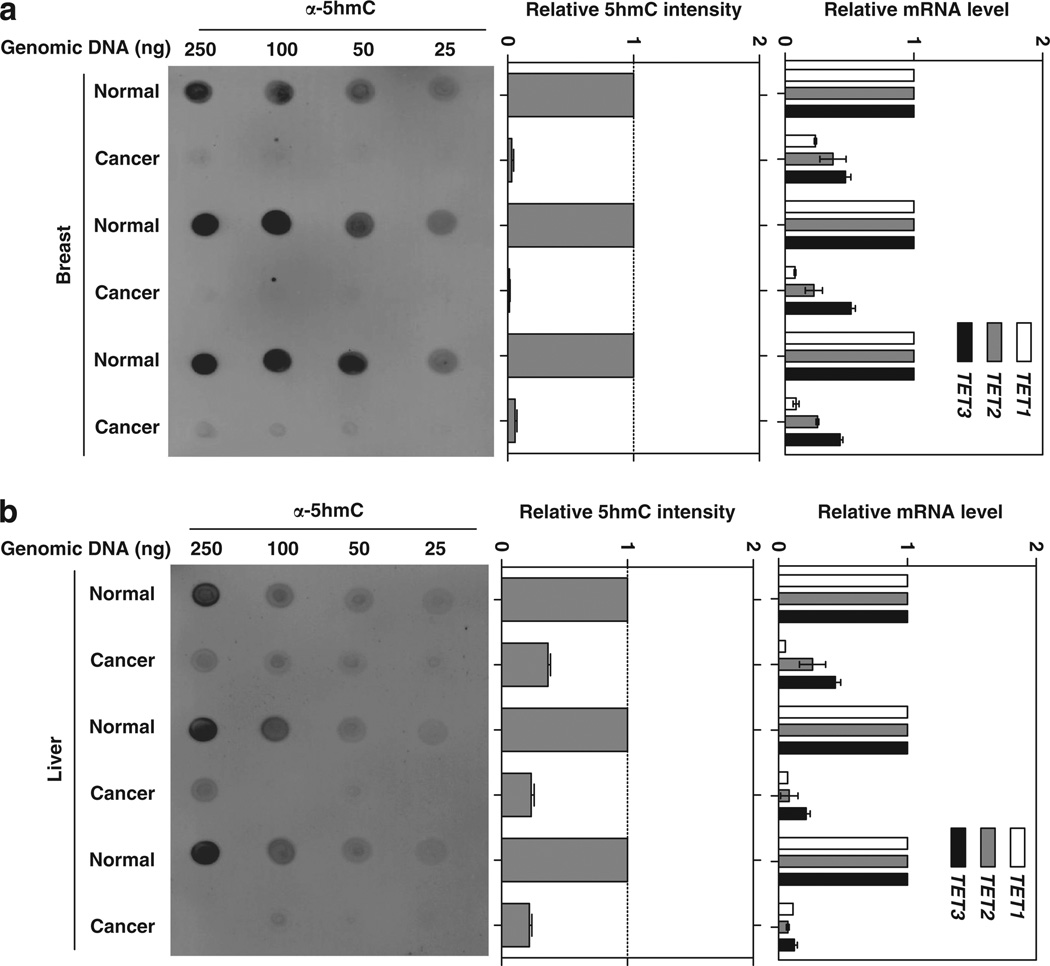
The TET (ten-eleven translocation) family of α-ketoglutarate (α-KG)-dependent dioxygenases catalyzes the sequential oxidation of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), 5-formylcytosine and 5-carboxylcytosine, leading to eventual DNA demethylation. The TET2 gene is a bona fide tumor suppressor frequently mutated in leukemia, and TET enzyme activity is inhibited in IDH1/2-mutated tumors by the oncometabolite 2-hydroxyglutarate, an antagonist of α-KG, linking 5mC oxidation to cancer development. We report here that the levels of 5hmC are dramatically reduced in human breast, liver, lung, pancreatic and prostate cancers when compared with the matched surrounding normal tissues. Associated with the 5hmC decrease is the substantial reduction of the expression of all three TET genes, revealing a possible mechanism for the reduced 5hmC in cancer cells. The decrease of 5hmC was also observed during tumor development in different genetically engineered mouse models. Together, our results identify 5hmC as a biomarker whose decrease is broadly and tightly associated with tumor development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Role of ten-eleven translocation proteins and 5-hydroxymethylcytosine in hepatocellular carcinoma.Cell Prolif. 2019 Jul;52(4):e12626. doi: 10.1111/cpr.12626. Epub 2019 Apr 29. Cell Prolif. 2019. PMID: 31033072 Free PMC article. Review.
-
Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine.Science. 2011 Sep 2;333(6047):1300-3. doi: 10.1126/science.1210597. Epub 2011 Jul 21. Science. 2011. PMID: 21778364 Free PMC article.
-
TET proteins and 5-methylcytosine oxidation in hematological cancers.Immunol Rev. 2015 Jan;263(1):6-21. doi: 10.1111/imr.12239. Immunol Rev. 2015. PMID: 25510268 Free PMC article. Review.
-
Connections between TET proteins and aberrant DNA modification in cancer.Trends Genet. 2014 Oct;30(10):464-74. doi: 10.1016/j.tig.2014.07.005. Epub 2014 Aug 14. Trends Genet. 2014. PMID: 25132561 Free PMC article. Review.
-
Distinct and overlapping control of 5-methylcytosine and 5-hydroxymethylcytosine by the TET proteins in human cancer cells.Genome Biol. 2014 Jun 23;15(6):R81. doi: 10.1186/gb-2014-15-6-r81. Genome Biol. 2014. PMID: 24958354 Free PMC article.
Cited by
-
TET1 upregulation drives cancer cell growth through aberrant enhancer hydroxymethylation of HMGA2 in hepatocellular carcinoma.Cancer Sci. 2021 Jul;112(7):2855-2869. doi: 10.1111/cas.14897. Epub 2021 May 10. Cancer Sci. 2021. PMID: 33970549 Free PMC article.
-
Loss of UHRF2 expression is associated with human neoplasia, promoter hypermethylation, decreased 5-hydroxymethylcytosine, and high proliferative activity.Oncotarget. 2016 Nov 15;7(46):76047-76061. doi: 10.18632/oncotarget.12583. Oncotarget. 2016. PMID: 27738314 Free PMC article.
-
ApcMin/+ tumours and normal mouse small intestines show linear metabolite concentration and DNA cytosine hydroxymethylation gradients from pylorus to colon.Sci Rep. 2020 Aug 12;10(1):13616. doi: 10.1038/s41598-020-70579-w. Sci Rep. 2020. PMID: 32788746 Free PMC article.
-
Molecular Tumor Subtypes of HPV-Positive Head and Neck Cancers: Biological Characteristics and Implications for Clinical Outcomes.Cancers (Basel). 2021 May 31;13(11):2721. doi: 10.3390/cancers13112721. Cancers (Basel). 2021. PMID: 34072836 Free PMC article. Review.
-
TET2 binds the androgen receptor and loss is associated with prostate cancer.Oncogene. 2017 Apr;36(15):2172-2183. doi: 10.1038/onc.2016.376. Epub 2016 Nov 7. Oncogene. 2017. PMID: 27819678 Free PMC article.
References
-
- Amary MF, Bacsi K, Maggiani F, Damato S, Halai D, Berisha F, et al. IDH1 and IDH2 mutations are frequent events in central chondrosarcoma and central and periosteal chondromas but not in other mesenchymal tumours. J Pathol. 2011a;224:334–343. - PubMed
-
- Amary MF, Damato S, Halai D, Eskandarpour M, Berisha F, Bonar F, et al. Ollier disease and Maffucci syndrome are caused by somatic mosaic mutations of IDH1 and IDH2. Nat Genet. 2011b;43:1262–1265. - PubMed
-
- Delhommeau F, Dupont S, Della VV, James C, Trannoy S, Masse A, et al. Mutation in TET2 in myeloid cancers. N Engl J Med. 2009;360:2289–2301. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
