

The MuvB complex sequentially recruits B-Myb and FoxM1 to promote mitotic gene expression
- PMID: 22391450
- PMCID: PMC3305985
- DOI: 10.1101/gad.181933.111
The MuvB complex sequentially recruits B-Myb and FoxM1 to promote mitotic gene expression
Abstract
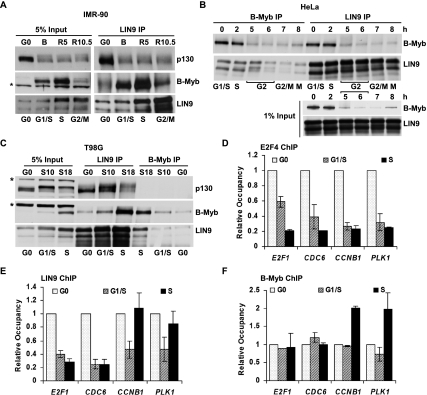
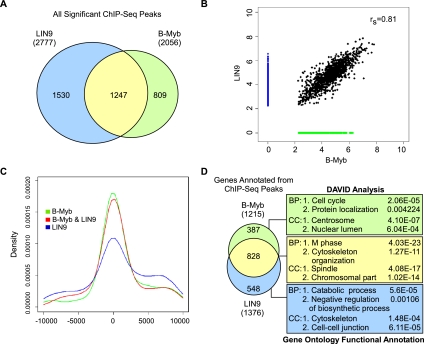
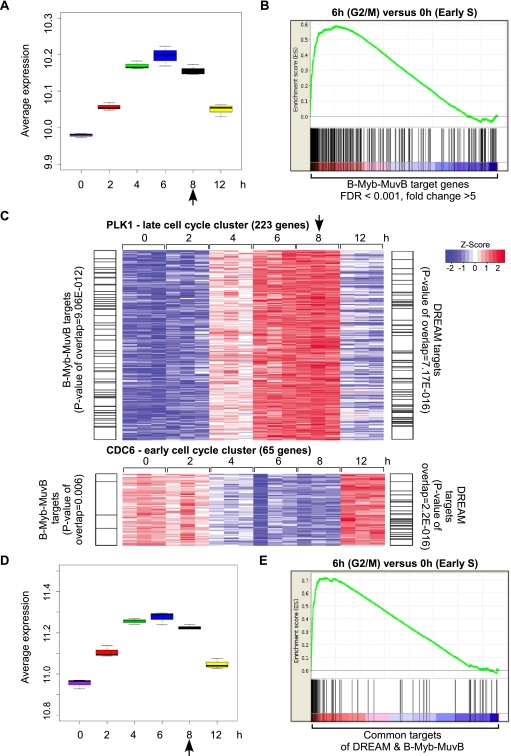
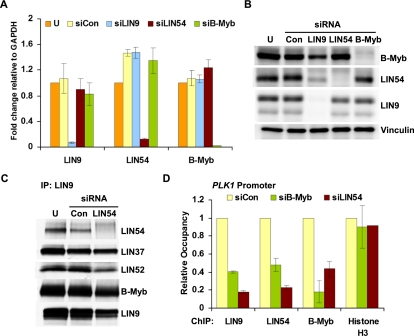
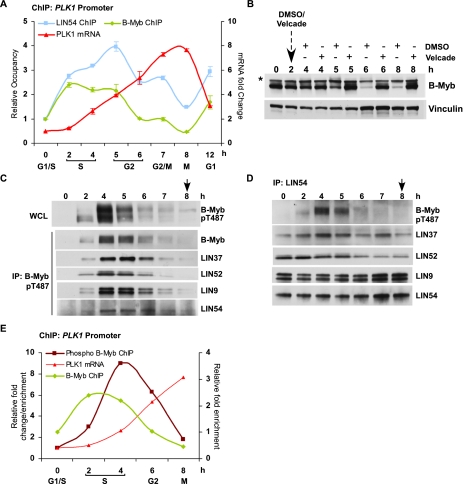
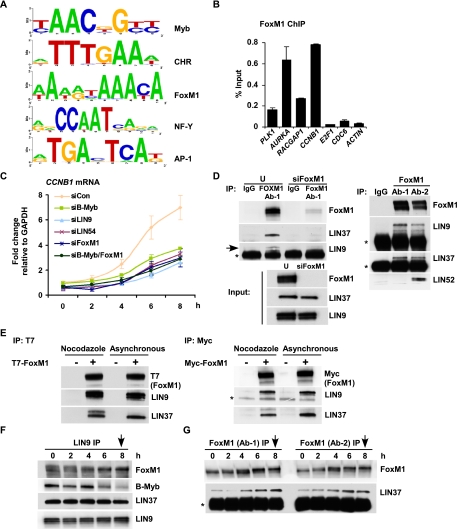
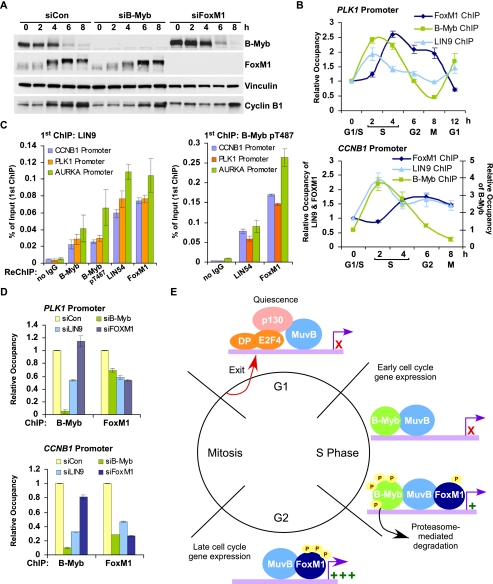
Cell cycle progression is dependent on two major waves of gene expression. Early cell cycle gene expression occurs during G1/S to generate factors required for DNA replication, while late cell cycle gene expression begins during G2 to prepare for mitosis. Here we demonstrate that the MuvB complex-comprised of LIN9, LIN37, LIN52, LIN54, and RBBP4-serves an essential role in three distinct transcription complexes to regulate cell cycle gene expression. The MuvB complex, together with the Rb-like protein p130, E2F4, and DP1, forms the DREAM complex during quiescence and represses expression of both early and late genes. Upon cell cycle entry, the MuvB complex dissociates from p130/DREAM, binds to B-Myb, and reassociates with the promoters of late genes during S phase. MuvB and B-Myb are required for the subsequent recruitment of FoxM1 to late gene promoters during G2. The MuvB complex remains bound to FoxM1 during peak late cell cycle gene expression, while B-Myb binding is lost when it undergoes phosphorylation-dependent, proteasome-mediated degradation during late S phase. Our results reveal a novel role for the MuvB complex in recruiting B-Myb and FoxM1 to promote late cell cycle gene expression and in regulating cell cycle gene expression from quiescence through mitosis.
Figures
Similar articles
-
The cell cycle regulatory DREAM complex is disrupted by high expression of oncogenic B-Myb.Oncogene. 2019 Feb;38(7):1080-1092. doi: 10.1038/s41388-018-0490-y. Epub 2018 Sep 11. Oncogene. 2019. PMID: 30206359 Free PMC article.
-
Simultaneous expression of MMB-FOXM1 complex components enables efficient bypass of senescence.Sci Rep. 2021 Nov 2;11(1):21506. doi: 10.1038/s41598-021-01012-z. Sci Rep. 2021. PMID: 34728711 Free PMC article.
-
Structural mechanisms of DREAM complex assembly and regulation.Genes Dev. 2015 May 1;29(9):961-74. doi: 10.1101/gad.257568.114. Epub 2015 Apr 27. Genes Dev. 2015. PMID: 25917549 Free PMC article.
-
MuvB: A Key to Cell Cycle Control in Ovarian Cancer.Front Oncol. 2018 Jun 11;8:223. doi: 10.3389/fonc.2018.00223. eCollection 2018. Front Oncol. 2018. PMID: 29942794 Free PMC article. Review.
-
Cell cycle transcription control: DREAM/MuvB and RB-E2F complexes.Crit Rev Biochem Mol Biol. 2017 Dec;52(6):638-662. doi: 10.1080/10409238.2017.1360836. Epub 2017 Aug 11. Crit Rev Biochem Mol Biol. 2017. PMID: 28799433 Review.
Cited by
-
MYBL2 and ATM suppress replication stress in pluripotent stem cells.EMBO Rep. 2021 May 5;22(5):e51120. doi: 10.15252/embr.202051120. Epub 2021 Mar 28. EMBO Rep. 2021. PMID: 33779025 Free PMC article.
-
Does Arabidopsis thaliana DREAM of cell cycle control?EMBO J. 2015 Aug 4;34(15):1987-9. doi: 10.15252/embj.201592196. Epub 2015 Jun 18. EMBO J. 2015. PMID: 26089020 Free PMC article.
-
An important role for Myb-MuvB and its target gene KIF23 in a mouse model of lung adenocarcinoma.Oncogene. 2017 Jan 5;36(1):110-121. doi: 10.1038/onc.2016.181. Epub 2016 May 23. Oncogene. 2017. PMID: 27212033
-
The role of FoxM1 in immune cells.Clin Exp Med. 2023 Oct;23(6):1973-1979. doi: 10.1007/s10238-023-01037-w. Epub 2023 Mar 13. Clin Exp Med. 2023. PMID: 36913035 Review.
-
Cooperation of cancer drivers with regulatory germline variants shapes clinical outcomes.Nat Commun. 2019 Sep 11;10(1):4128. doi: 10.1038/s41467-019-12071-2. Nat Commun. 2019. PMID: 31511524 Free PMC article.
References
-
- Breeden LL 2003. Periodic transcription: a cycle within a cycle. Curr Biol 13: R31–R38 doi: 10.1016/S0960-9822(02)01386-6 - PubMed
-
- Caretti G, Salsi V, Vecchi C, Imbriano C, Mantovani R 2003. Dynamic recruitment of NF-Y and histone acetyltransferases on cell-cycle promoters. J Biol Chem 278: 30435–30440 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous