

Fibroblast growth factor 22 contributes to the development of retinal nerve terminals in the dorsal lateral geniculate nucleus
- PMID: 22363257
- PMCID: PMC3306139
- DOI: 10.3389/fnmol.2011.00061
Fibroblast growth factor 22 contributes to the development of retinal nerve terminals in the dorsal lateral geniculate nucleus
Abstract
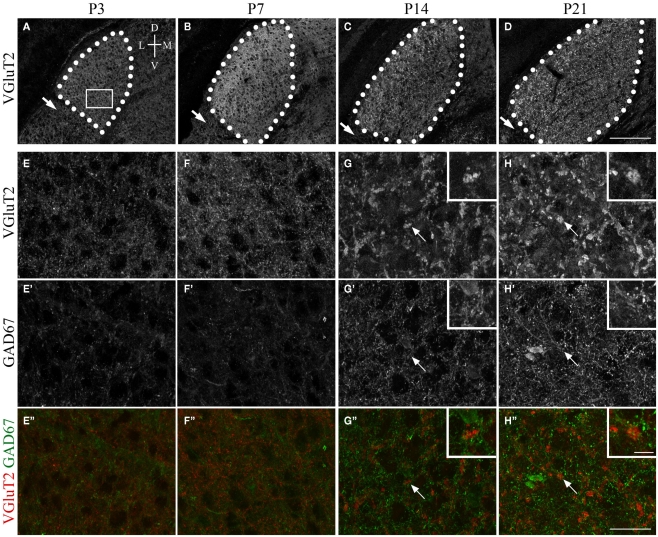
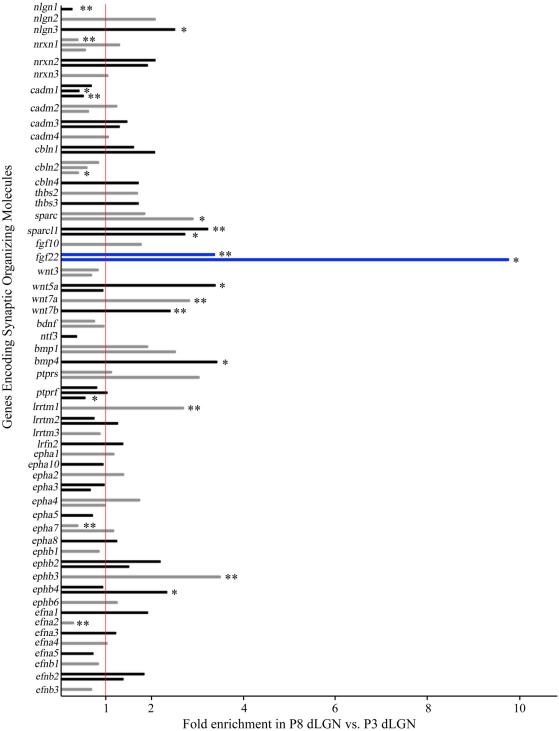
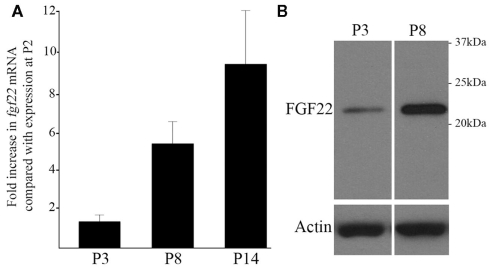
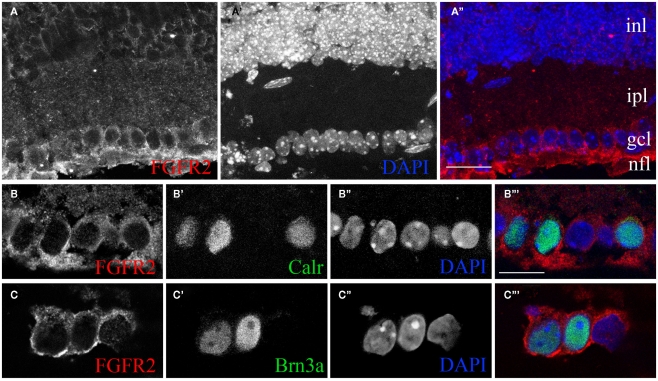
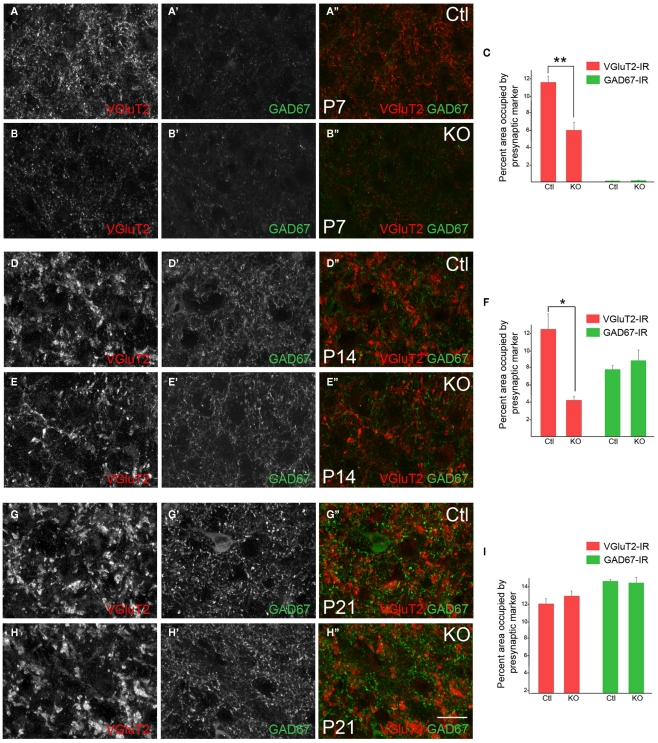
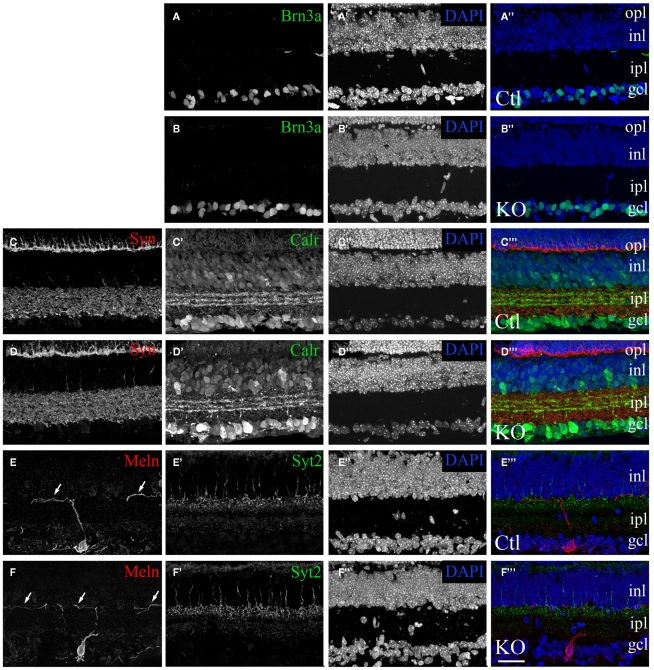
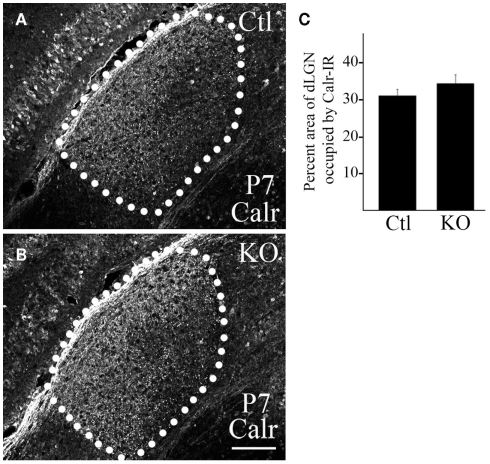
At least three forms of signaling between pre- and postsynaptic partners are necessary during synapse formation. First, "targeting" signals instruct presynaptic axons to recognize and adhere to the correct portion of a postsynaptic target cell. Second, trans-synaptic "organizing" signals induce differentiation in their synaptic partner so that each side of the synapse is specialized for synaptic transmission. Finally, in many regions of the nervous system an excess of synapses are initially formed, therefore "refinement" signals must either stabilize or destabilize the synapse to reinforce or eliminate connections, respectively. Because of both their importance in processing visual information and their accessibility, retinogeniculate synapses have served as a model for studying synaptic development. Molecular signals that drive retinogeniculate "targeting" and "refinement" have been identified, however, little is known about what "organizing" cues are necessary for the differentiation of retinal axons into presynaptic terminals. To identify such "organizing" cues, we used microarray analysis to assess whether any target-derived "synaptic organizers" were enriched in the mouse dorsal lateral geniculate nucleus (dLGN) during retinogeniculate synapse formation. One candidate "organizing" molecule enriched in perinatal dLGN was FGF22, a secreted cue that induces the formation of excitatory nerve terminals in muscle, hippocampus, and cerebellum. In FGF22 knockout mice, the development of retinal terminals in dLGN was impaired. Thus, FGF22 is an important "organizing" cue for the timely development of retinogeniculate synapses.
Figures
Similar articles
-
Nuclei-specific differences in nerve terminal distribution, morphology, and development in mouse visual thalamus.Neural Dev. 2014 Jul 10;9:16. doi: 10.1186/1749-8104-9-16. Neural Dev. 2014. PMID: 25011644 Free PMC article.
-
Synaptic development of the mouse dorsal lateral geniculate nucleus.J Comp Neurol. 2010 Mar 1;518(5):622-35. doi: 10.1002/cne.22223. J Comp Neurol. 2010. PMID: 20034053 Free PMC article.
-
Distinct FGFs promote differentiation of excitatory and inhibitory synapses.Nature. 2010 Jun 10;465(7299):783-7. doi: 10.1038/nature09041. Epub 2010 May 26. Nature. 2010. PMID: 20505669 Free PMC article.
-
An evolving view of retinogeniculate transmission.Vis Neurosci. 2017 Jan;34:E013. doi: 10.1017/S0952523817000104. Vis Neurosci. 2017. PMID: 28965513 Free PMC article. Review.
-
Organization, Function, and Development of the Mouse Retinogeniculate Synapse.Annu Rev Vis Sci. 2020 Sep 15;6:261-285. doi: 10.1146/annurev-vision-121219-081753. Annu Rev Vis Sci. 2020. PMID: 32936733 Review.
Cited by
-
Retinal input regulates the timing of corticogeniculate innervation.J Neurosci. 2013 Jun 12;33(24):10085-97. doi: 10.1523/JNEUROSCI.5271-12.2013. J Neurosci. 2013. PMID: 23761904 Free PMC article.
-
A molecular mechanism regulating the timing of corticogeniculate innervation.Cell Rep. 2013 Nov 14;5(3):573-81. doi: 10.1016/j.celrep.2013.09.041. Epub 2013 Oct 31. Cell Rep. 2013. PMID: 24183669 Free PMC article.
-
FGF22 signaling regulates synapse formation during post-injury remodeling of the spinal cord.EMBO J. 2015 May 5;34(9):1231-43. doi: 10.15252/embj.201490578. Epub 2015 Mar 12. EMBO J. 2015. PMID: 25766255 Free PMC article.
-
Development of astrocyte morphology and function in mouse visual thalamus.J Comp Neurol. 2022 May;530(7):945-962. doi: 10.1002/cne.25261. Epub 2021 Oct 25. J Comp Neurol. 2022. PMID: 34636034 Free PMC article.
-
Selective Inactivation of Fibroblast Growth Factor 22 (FGF22) in CA3 Pyramidal Neurons Impairs Local Synaptogenesis and Affective Behavior Without Affecting Dentate Neurogenesis.Front Synaptic Neurosci. 2017 Dec 19;9:17. doi: 10.3389/fnsyn.2017.00017. eCollection 2017. Front Synaptic Neurosci. 2017. PMID: 29311892 Free PMC article.
References
-
- Bjartmar L., Huberman A. D., Ullian E. M., Renteria R. C., Liu X., Xu W., Prezioso J., Susman M. W., Stellwagen D., Stokes C. C., Cho R., Worley P., Malenka R. C., Ball S., Peachey N. S., Copenhagen D., Chapman B., Nakamoto M., Barres B. A., Perin M. S. (2006). Neuronal pentraxins mediate synaptic refinement in the developing visual system. J. Neurosci. 26, 6269–628110.1523/JNEUROSCI.4212-05.2006 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases