

The splicing regulator Rbfox2 is required for both cerebellar development and mature motor function
- PMID: 22357600
- PMCID: PMC3305983
- DOI: 10.1101/gad.182477.111
The splicing regulator Rbfox2 is required for both cerebellar development and mature motor function
Abstract
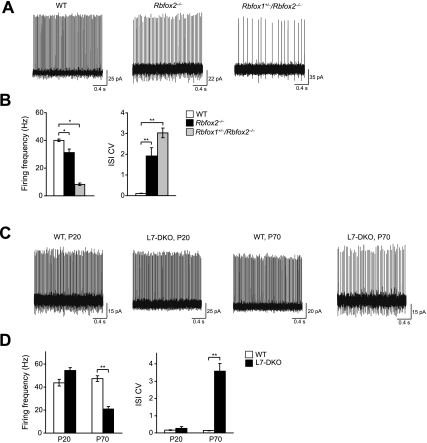
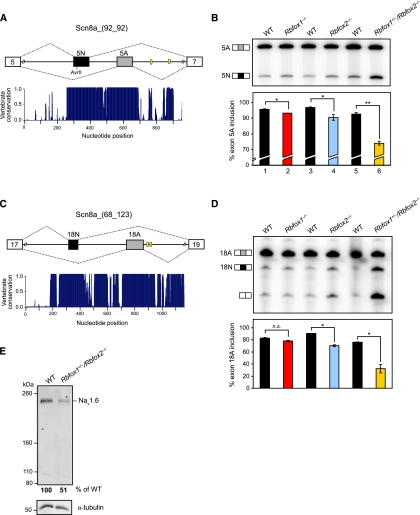
The Rbfox proteins (Rbfox1, Rbfox2, and Rbfox3) regulate the alternative splicing of many important neuronal transcripts and have been implicated in a variety of neurological disorders. However, their roles in brain development and function are not well understood, in part due to redundancy in their activities. Here we show that, unlike Rbfox1 deletion, the CNS-specific deletion of Rbfox2 disrupts cerebellar development. Genome-wide analysis of Rbfox2(-/-) brain RNA identifies numerous splicing changes altering proteins important both for brain development and mature neuronal function. To separate developmental defects from alterations in the physiology of mature cells, Rbfox1 and Rbfox2 were deleted from mature Purkinje cells, resulting in highly irregular firing. Notably, the Scn8a mRNA encoding the Na(v)1.6 sodium channel, a key mediator of Purkinje cell pacemaking, is improperly spliced in RbFox2 and Rbfox1 mutant brains, leading to highly reduced protein expression. Thus, Rbfox2 protein controls a post-transcriptional program required for proper brain development. Rbfox2 is subsequently required with Rbfox1 to maintain mature neuronal physiology, specifically Purkinje cell pacemaking, through their shared control of sodium channel transcript splicing.
Figures
Similar articles
-
Expression of RNA-binding protein Rbfox1l demarcates a restricted population of dorsal telencephalic neurons within the adult zebrafish brain.Gene Expr Patterns. 2019 Jan;31:32-41. doi: 10.1016/j.gep.2019.01.001. Epub 2019 Jan 8. Gene Expr Patterns. 2019. PMID: 30634066
-
Rbfox proteins regulate alternative splicing of neuronal sodium channel SCN8A.Mol Cell Neurosci. 2012 Feb;49(2):120-6. doi: 10.1016/j.mcn.2011.10.005. Epub 2011 Oct 21. Mol Cell Neurosci. 2012. PMID: 22044765 Free PMC article.
-
The RNA-binding protein Rbfox2: an essential regulator of EMT-driven alternative splicing and a mediator of cellular invasion.Oncogene. 2014 Feb 27;33(9):1082-92. doi: 10.1038/onc.2013.50. Epub 2013 Feb 25. Oncogene. 2014. PMID: 23435423
-
Developmental regulation of RNA processing by Rbfox proteins.Wiley Interdiscip Rev RNA. 2017 Mar;8(2):10.1002/wrna.1398. doi: 10.1002/wrna.1398. Epub 2016 Oct 17. Wiley Interdiscip Rev RNA. 2017. PMID: 27748060 Free PMC article. Review.
-
RBFOX2 protein domains and cellular activities.Biochem Soc Trans. 2014 Aug;42(4):1180-3. doi: 10.1042/BST20140050. Biochem Soc Trans. 2014. PMID: 25110022 Review.
Cited by
-
RNA processing in neurological tissue: development, aging and disease.Semin Cell Dev Biol. 2021 Jun;114:57-67. doi: 10.1016/j.semcdb.2020.09.004. Epub 2020 Oct 16. Semin Cell Dev Biol. 2021. PMID: 33077405 Free PMC article. Review.
-
mRNA Processing: An Emerging Frontier in the Regulation of Pancreatic β Cell Function.Front Genet. 2020 Sep 1;11:983. doi: 10.3389/fgene.2020.00983. eCollection 2020. Front Genet. 2020. PMID: 33088281 Free PMC article. Review.
-
Zika Virus Neuropathogenesis: The Different Brain Cells, Host Factors and Mechanisms Involved.Front Immunol. 2022 Mar 16;13:773191. doi: 10.3389/fimmu.2022.773191. eCollection 2022. Front Immunol. 2022. PMID: 35371036 Free PMC article. Review.
-
Novel Rbfox2 isoforms associated with alternative exon usage in rat cortex and suprachiasmatic nucleus.Sci Rep. 2017 Aug 30;7(1):9929. doi: 10.1038/s41598-017-10535-3. Sci Rep. 2017. PMID: 28855650 Free PMC article.
-
RBFOX3/NeuN is Required for Hippocampal Circuit Balance and Function.Sci Rep. 2015 Dec 1;5:17383. doi: 10.1038/srep17383. Sci Rep. 2015. PMID: 26619789 Free PMC article.
References
-
- Barski JJ, Dethleffsen K, Meyer M 2000. Cre recombinase expression in cerebellar Purkinje cells. Genesis 28: 93–98 - PubMed
-
- Bhalla K, Phillips HA, Crawford J, McKenzie OL, Mulley JC, Eyre H, Gardner AE, Kremmidiotis G, Callen DF 2004. The de novo chromosome 16 translocations of two patients with abnormal phenotypes (mental retardation and epilepsy) disrupt the A2BP1 gene. J Hum Genet 49: 308–311 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases