

Cytoplasmic Arabidopsis AGO7 accumulates in membrane-associated siRNA bodies and is required for ta-siRNA biogenesis
- PMID: 22327216
- PMCID: PMC3321200
- DOI: 10.1038/emboj.2012.20
Cytoplasmic Arabidopsis AGO7 accumulates in membrane-associated siRNA bodies and is required for ta-siRNA biogenesis
Abstract
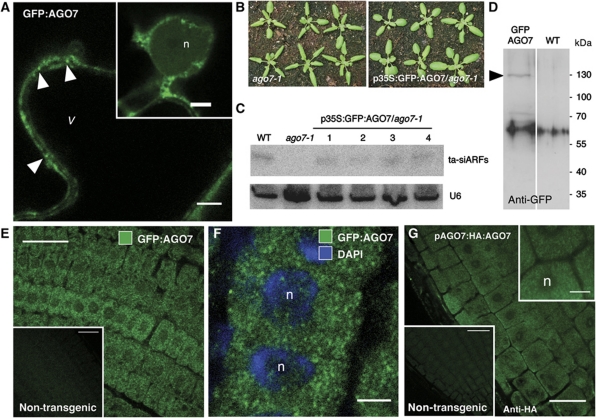
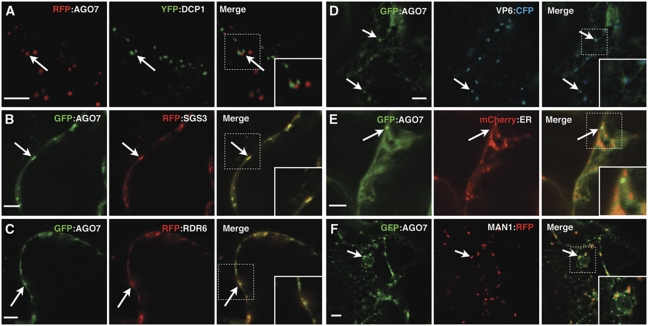
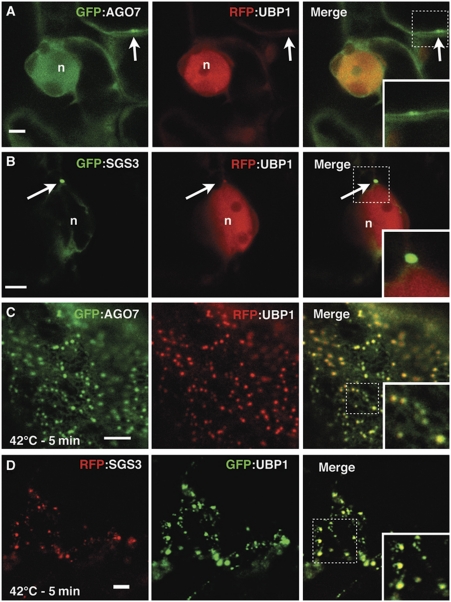
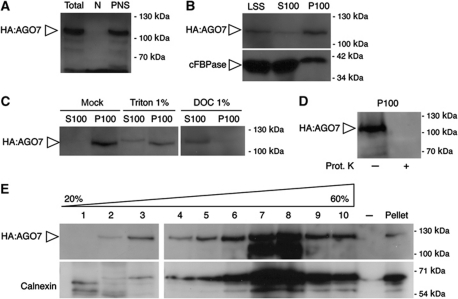
Formation of trans-acting small interfering RNAs (ta-siRNAs) from the TAS3 precursor is triggered by the AGO7/miR390 complex, which primes TAS3 for conversion into double-stranded RNA by the RNA-dependent RNA polymerase RDR6 and SGS3. These ta-siRNAs control several aspects of plant development. The mechanism routing AGO7-cleaved TAS3 precursor to RDR6/SGS3 and its subcellular organization are unknown. We show that AGO7 accumulates together with SGS3 and RDR6 in cytoplasmic siRNA bodies that are distinct from P-bodies. siRNA bodies colocalize with a membrane-associated viral protein and become positive for stress-granule markers upon stress-induced translational repression, this suggests that siRNA bodies are membrane-associated sites of accumulation of mRNA stalled during translation. AGO7 congregates with miR390 and SGS3 in membranes and its targeting to the nucleus prevents its accumulation in siRNA bodies and ta-siRNA formation. AGO7 is therefore required in the cytoplasm and membranous siRNA bodies for TAS3 processing, revealing a hitherto unknown role for membrane-associated ribonucleoparticles in ta-siRNA biogenesis and AGO action in plants.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7.Curr Biol. 2006 May 9;16(9):927-32. doi: 10.1016/j.cub.2006.03.035. Curr Biol. 2006. PMID: 16682354
-
Cooperative recruitment of RDR6 by SGS3 and SDE5 during small interfering RNA amplification in Arabidopsis.Proc Natl Acad Sci U S A. 2021 Aug 24;118(34):e2102885118. doi: 10.1073/pnas.2102885118. Proc Natl Acad Sci U S A. 2021. PMID: 34408020 Free PMC article.
-
RDR6-mediated synthesis of complementary RNA is terminated by miRNA stably bound to template RNA.Nucleic Acids Res. 2012 Jan;40(2):594-9. doi: 10.1093/nar/gkr760. Epub 2011 Sep 19. Nucleic Acids Res. 2012. PMID: 21930511 Free PMC article.
-
MicroRNA-dependent trans-acting siRNA production.Sci STKE. 2005 Sep 6;2005(300):pe43. doi: 10.1126/stke.3002005pe43. Sci STKE. 2005. PMID: 16145017 Review.
-
miRNAs in the biogenesis of trans-acting siRNAs in higher plants.Semin Cell Dev Biol. 2010 Oct;21(8):798-804. doi: 10.1016/j.semcdb.2010.03.008. Epub 2010 Mar 30. Semin Cell Dev Biol. 2010. PMID: 20359543 Review.
Cited by
-
Lessons on RNA silencing mechanisms in plants from eukaryotic argonaute structures.Plant Cell. 2013 Jan;25(1):22-37. doi: 10.1105/tpc.112.105643. Epub 2013 Jan 9. Plant Cell. 2013. PMID: 23303917 Free PMC article.
-
In plants, decapping prevents RDR6-dependent production of small interfering RNAs from endogenous mRNAs.Nucleic Acids Res. 2015 Mar 11;43(5):2902-13. doi: 10.1093/nar/gkv119. Epub 2015 Feb 18. Nucleic Acids Res. 2015. PMID: 25694514 Free PMC article.
-
Plant and animal small RNA communications between cells and organisms.Nat Rev Mol Cell Biol. 2022 Mar;23(3):185-203. doi: 10.1038/s41580-021-00425-y. Epub 2021 Oct 27. Nat Rev Mol Cell Biol. 2022. PMID: 34707241 Free PMC article. Review.
-
The plant siRNA landscape.Plant Cell. 2024 Jan 30;36(2):246-275. doi: 10.1093/plcell/koad253. Plant Cell. 2024. PMID: 37772967 Free PMC article. Review.
-
Biogenesis of phased siRNAs on membrane-bound polysomes in Arabidopsis.Elife. 2016 Dec 12;5:e22750. doi: 10.7554/eLife.22750. Elife. 2016. PMID: 27938667 Free PMC article.
References
-
- Adenot X, Elmayan T, Lauressergues D, Boutet S, Bouche N, Gasciolli V, Vaucheret H (2006) DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Curr Biol 16: 927–932 - PubMed
-
- Allen E, Xie Z, Gustafson AM, Carrington JC (2005) microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121: 207–221 - PubMed
-
- Anderson P, Kedersha N (2009) RNA granules: post-transcriptional and epigenetic modulators of gene expression. Nat Rev Mol Cell Biol 10: 430–436 - PubMed
-
- Axtell MJ, Jan C, Rajagopalan R, Bartel DP (2006) A two-hit trigger for siRNA biogenesis in plants. Cell 127: 565–577 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
