

The vital role of polymerase ζ and REV1 in mutagenic, but not correct, DNA synthesis across benzo[a]pyrene-dG and recruitment of polymerase ζ by REV1 to replication-stalled site
- PMID: 22303021
- PMCID: PMC3308759
- DOI: 10.1074/jbc.M111.331728
The vital role of polymerase ζ and REV1 in mutagenic, but not correct, DNA synthesis across benzo[a]pyrene-dG and recruitment of polymerase ζ by REV1 to replication-stalled site
Abstract
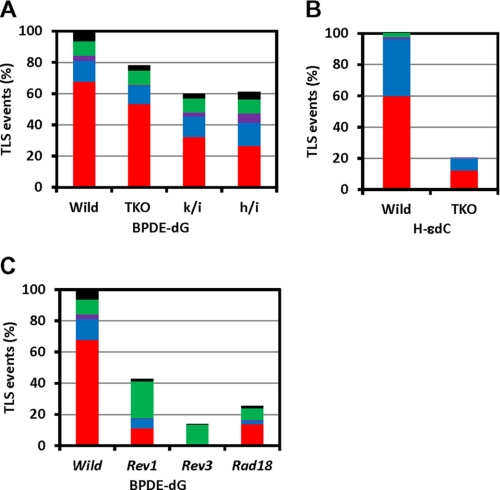
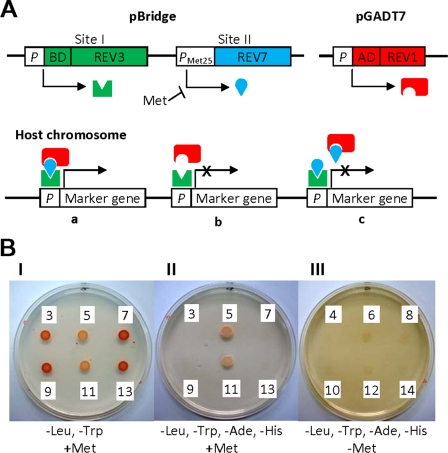
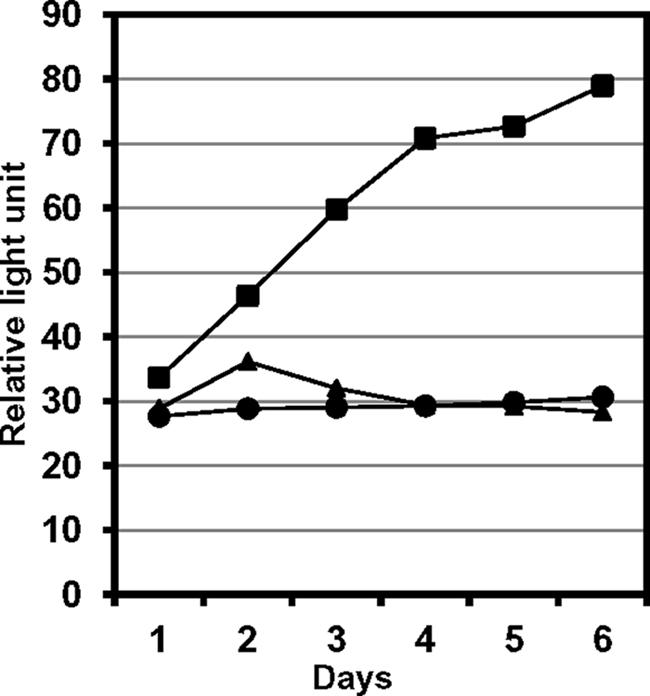
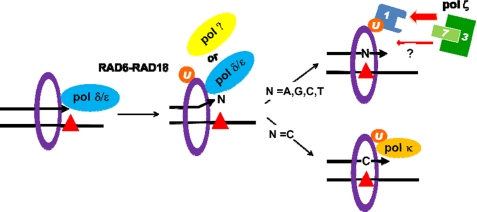
The DNA synthesis across DNA lesions, termed translesion synthesis (TLS), is a complex process influenced by various factors. To investigate this process in mammalian cells, we examined TLS across a benzo[a]pyrene dihydrodiol epoxide-derived dG adduct (BPDE-dG) using a plasmid bearing a single BPDE-dG and genetically engineered mouse embryonic fibroblasts (MEFs). In wild-type MEFs, TLS was extremely miscoding (>90%) with G → T transversions being predominant. Knockout of the Rev1 gene decreased both the TLS efficiency and the miscoding frequency. Knockout of the Rev3L gene, coding for the catalytic subunit of pol ζ, caused even greater decreases in these two TLS parameters; almost all residual TLS were error-free. Thus, REV1 and pol ζ are critical to mutagenic, but not accurate, TLS across BPDE-dG. The introduction of human REV1 cDNA into Rev1(-/-) MEFs restored the mutagenic TLS, but a REV1 mutant lacking the C terminus did not. Yeast and mammalian three-hybrid assays revealed that the REV7 subunit of pol ζ mediated the interaction between REV3 and the REV1 C terminus. These results support the hypothesis that REV1 recruits pol ζ through the interaction with REV7. Our results also predict the existence of a minor REV1-independent pol ζ recruitment pathway. Finally, although mutagenic TLS across BPDE-dG largely depends on RAD18, experiments using Polk(-/-) Polh(-/-) Poli(-/-) triple-gene knockout MEFs unexpectedly revealed that another polymerase(s) could insert a nucleotide opposite BPDE-dG. This indicates that a non-Y family polymerase(s) can insert a nucleotide opposite BPDE-dG, but the subsequent extension from miscoding termini depends on REV1-polζ in a RAD18-dependent manner.
Figures
Similar articles
-
Translesion Synthesis of the N(2)-2'-Deoxyguanosine Adduct of the Dietary Mutagen IQ in Human Cells: Error-Free Replication by DNA Polymerase κ and Mutagenic Bypass by DNA Polymerases η, ζ, and Rev1.Chem Res Toxicol. 2016 Sep 19;29(9):1549-59. doi: 10.1021/acs.chemrestox.6b00221. Epub 2016 Aug 17. Chem Res Toxicol. 2016. PMID: 27490094 Free PMC article.
-
Poleta, Polzeta and Rev1 together are required for G to T transversion mutations induced by the (+)- and (-)-trans-anti-BPDE-N2-dG DNA adducts in yeast cells.Nucleic Acids Res. 2006 Jan 13;34(2):417-25. doi: 10.1093/nar/gkj446. Print 2006. Nucleic Acids Res. 2006. PMID: 16415180 Free PMC article.
-
Crystal structure of human REV7 in complex with a human REV3 fragment and structural implication of the interaction between DNA polymerase zeta and REV1.J Biol Chem. 2010 Apr 16;285(16):12299-307. doi: 10.1074/jbc.M109.092403. Epub 2010 Feb 17. J Biol Chem. 2010. PMID: 20164194 Free PMC article.
-
Roles of mutagenic translesion synthesis in mammalian genome stability, health and disease.DNA Repair (Amst). 2015 May;29:56-64. doi: 10.1016/j.dnarep.2015.01.001. Epub 2015 Jan 21. DNA Repair (Amst). 2015. PMID: 25655219 Review.
-
DNA polymerase zeta: new insight into eukaryotic mutagenesis and mammalian embryonic development.World J Gastroenterol. 2003 Jun;9(6):1165-9. doi: 10.3748/wjg.v9.i6.1165. World J Gastroenterol. 2003. PMID: 12800216 Free PMC article. Review.
Cited by
-
Preferred WMSA catalytic mechanism of the nucleotidyl transfer reaction in human DNA polymerase κ elucidates error-free bypass of a bulky DNA lesion.Nucleic Acids Res. 2012 Oct;40(18):9193-205. doi: 10.1093/nar/gks653. Epub 2012 Jul 5. Nucleic Acids Res. 2012. PMID: 22772988 Free PMC article.
-
Within-Host Variations of Human Papillomavirus Reveal APOBEC Signature Mutagenesis in the Viral Genome.J Virol. 2018 May 29;92(12):e00017-18. doi: 10.1128/JVI.00017-18. Print 2018 Jun 15. J Virol. 2018. PMID: 29593040 Free PMC article.
-
Targeting REV7 effectively reverses 5-FU and oxaliplatin resistance in colorectal cancer.Cancer Cell Int. 2020 Dec 3;20(1):580. doi: 10.1186/s12935-020-01668-z. Cancer Cell Int. 2020. PMID: 33292253 Free PMC article.
-
Y-family DNA polymerase-independent gap-filling translesion synthesis across aristolochic acid-derived adenine adducts in mouse cells.DNA Repair (Amst). 2016 Oct;46:55-60. doi: 10.1016/j.dnarep.2016.07.003. Epub 2016 Jul 29. DNA Repair (Amst). 2016. PMID: 27497692 Free PMC article.
-
Human Pol ζ purified with accessory subunits is active in translesion DNA synthesis and complements Pol η in cisplatin bypass.Proc Natl Acad Sci U S A. 2014 Feb 25;111(8):2954-9. doi: 10.1073/pnas.1324001111. Epub 2014 Jan 21. Proc Natl Acad Sci U S A. 2014. PMID: 24449906 Free PMC article.
References
-
- Ohmori H., Friedberg E. C., Fuchs R. P., Goodman M. F., Hanaoka F., Hinkle D., Kunkel T. A., Lawrence C. W., Livneh Z., Nohmi T., Prakash L., Prakash S., Todo T., Walker G. C., Wang Z., Woodgate R. (2001) The Y-family of DNA polymerases. Mol. Cell 8, 7–8 - PubMed
-
- Masutani C., Kusumoto R., Yamada A., Dohmae N., Yokoi M., Yuasa M., Araki M., Iwai S., Takio K., Hanaoka F. (1999) The XPV (xeroderma pigmentosum variant) gene encodes human DNA polymerase ϵ. Nature 399, 700–704 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
