

Amyotrophic lateral sclerosis-associated mutant VAPBP56S perturbs calcium homeostasis to disrupt axonal transport of mitochondria
- PMID: 22258555
- PMCID: PMC3315205
- DOI: 10.1093/hmg/dds011
Amyotrophic lateral sclerosis-associated mutant VAPBP56S perturbs calcium homeostasis to disrupt axonal transport of mitochondria
Abstract
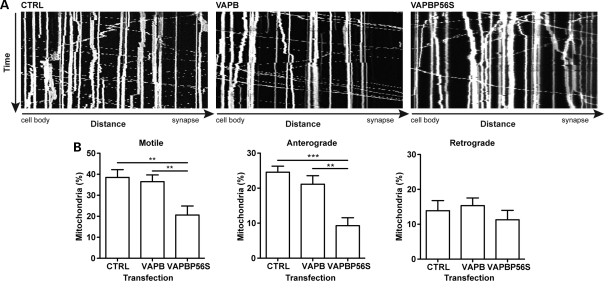
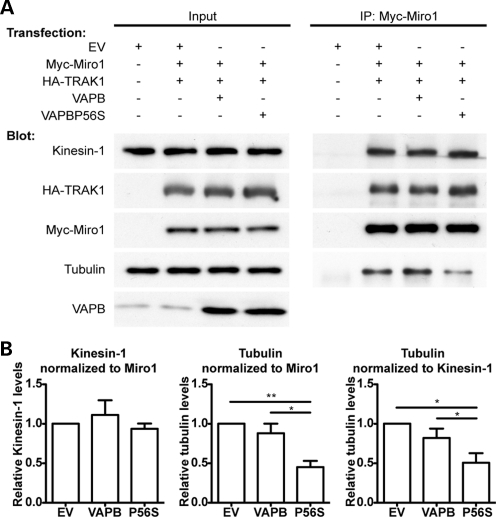
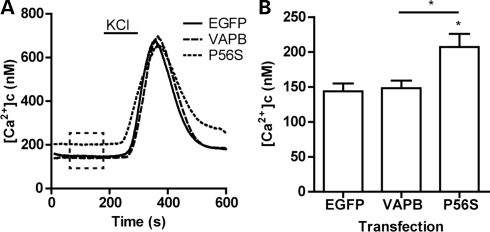
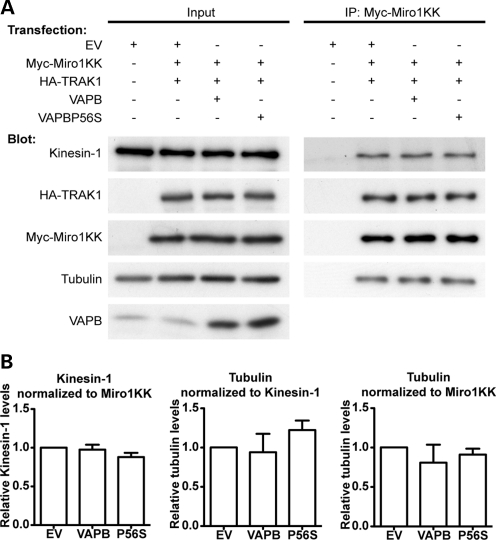
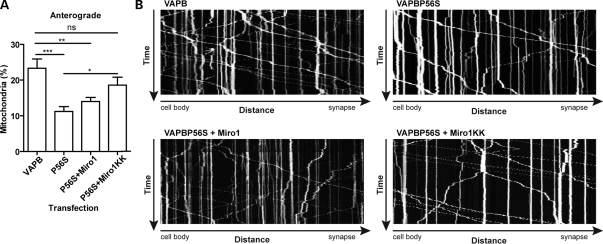
A proline-to-serine substitution at position 56 in the gene encoding vesicle-associated membrane protein-associated protein B (VAPB; VAPBP56S) causes some dominantly inherited familial forms of motor neuron disease, including amyotrophic lateral sclerosis (ALS) type-8. Here, we show that expression of ALS mutant VAPBP56S but not wild-type VAPB in neurons selectively disrupts anterograde axonal transport of mitochondria. VAPBP56S-induced disruption of mitochondrial transport involved reductions in the frequency, velocity and persistence of anterograde mitochondrial movement. Anterograde axonal transport of mitochondria is mediated by the microtubule-based molecular motor kinesin-1. Attachment of kinesin-1 to mitochondria involves the outer mitochondrial membrane protein mitochondrial Rho GTPase-1 (Miro1) which acts as a sensor for cytosolic calcium levels ([Ca(2+)]c); elevated [Ca(2+)]c disrupts mitochondrial transport via an effect on Miro1. To gain insight into the mechanisms underlying the VAPBP56S effect on mitochondrial transport, we monitored [Ca(2+)]c levels in VAPBP56S-expressing neurons. Expression of VAPBP56S but not VAPB increased resting [Ca(2+)]c and this was associated with a reduction in the amounts of tubulin but not kinesin-1 that were associated with Miro1. Moreover, expression of a Ca(2+) insensitive mutant of Miro1 rescued defective mitochondrial axonal transport and restored the amounts of tubulin associated with the Miro1/kinesin-1 complex to normal in VAPBP56S-expressing cells. Our results suggest that ALS mutant VAPBP56S perturbs anterograde mitochondrial axonal transport by disrupting Ca(2+) homeostasis and effecting the Miro1/kinesin-1 interaction with tubulin.
Figures
Similar articles
-
Amyotrophic lateral sclerosis-associated mutant SOD1 inhibits anterograde axonal transport of mitochondria by reducing Miro1 levels.Hum Mol Genet. 2017 Dec 1;26(23):4668-4679. doi: 10.1093/hmg/ddx348. Hum Mol Genet. 2017. PMID: 28973175 Free PMC article.
-
VAPB interacts with the mitochondrial protein PTPIP51 to regulate calcium homeostasis.Hum Mol Genet. 2012 Mar 15;21(6):1299-311. doi: 10.1093/hmg/ddr559. Epub 2011 Nov 30. Hum Mol Genet. 2012. PMID: 22131369 Free PMC article.
-
Loss of Miro1-directed mitochondrial movement results in a novel murine model for neuron disease.Proc Natl Acad Sci U S A. 2014 Sep 2;111(35):E3631-40. doi: 10.1073/pnas.1402449111. Epub 2014 Aug 18. Proc Natl Acad Sci U S A. 2014. PMID: 25136135 Free PMC article.
-
Miro (Mitochondrial Rho GTPase), a key player of mitochondrial axonal transport and mitochondrial dynamics in neurodegenerative diseases.Mitochondrion. 2021 Jan;56:118-135. doi: 10.1016/j.mito.2020.10.005. Epub 2020 Oct 28. Mitochondrion. 2021. PMID: 33127590 Review.
-
The axonal transport of mitochondria.J Cell Sci. 2012 May 1;125(Pt 9):2095-104. doi: 10.1242/jcs.053850. Epub 2012 May 22. J Cell Sci. 2012. PMID: 22619228 Free PMC article. Review.
Cited by
-
Tobacco Smoke Exposure Exacerbated Crystalline Silica-Induced Lung Toxicity in Rats.Toxicol Sci. 2020 Dec 1;178(2):375-390. doi: 10.1093/toxsci/kfaa146. Toxicol Sci. 2020. PMID: 32976597 Free PMC article.
-
Sensory neuropathy-causing mutations in ATL3 affect ER-mitochondria contact sites and impair axonal mitochondrial distribution.Hum Mol Genet. 2019 Feb 15;28(4):615-627. doi: 10.1093/hmg/ddy352. Hum Mol Genet. 2019. PMID: 30339187 Free PMC article.
-
Whole blood transcriptome analysis in amyotrophic lateral sclerosis: A biomarker study.PLoS One. 2018 Jun 25;13(6):e0198874. doi: 10.1371/journal.pone.0198874. eCollection 2018. PLoS One. 2018. PMID: 29939990 Free PMC article.
-
Mitochondrial transport serves as a mitochondrial quality control strategy in axons: Implications for central nervous system disorders.CNS Neurosci Ther. 2019 Jul;25(7):876-886. doi: 10.1111/cns.13122. Epub 2019 Mar 21. CNS Neurosci Ther. 2019. PMID: 30900394 Free PMC article. Review.
-
Endoplasmic reticulum and mitochondria in diseases of motor and sensory neurons: a broken relationship?Cell Death Dis. 2018 Feb 28;9(3):333. doi: 10.1038/s41419-017-0125-1. Cell Death Dis. 2018. PMID: 29491369 Free PMC article. Review.
References
-
- Tanaka Y., Kanai Y., Okada Y., Nonaka S., Takeda S., Harada A., Hirokawa N. Targeted disruption of mouse conventional kinesin heavy chain, kif5B, results in abnormal perinuclear clustering of mitochondria. Cell. 1998;93:1147–1158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
