

Rational design of vaccines to elicit broadly neutralizing antibodies to HIV-1
- PMID: 22229123
- PMCID: PMC3234457
- DOI: 10.1101/cshperspect.a007278
Rational design of vaccines to elicit broadly neutralizing antibodies to HIV-1
Abstract
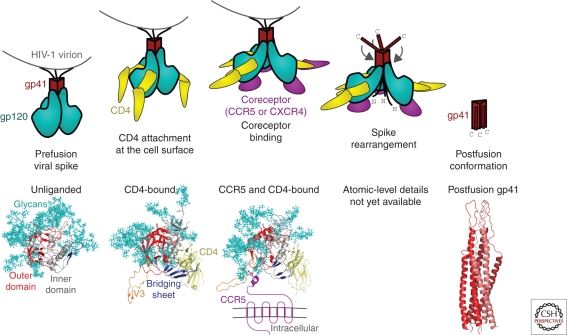
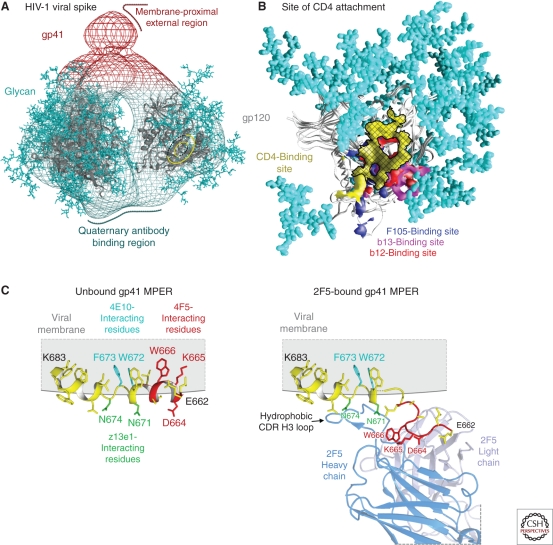
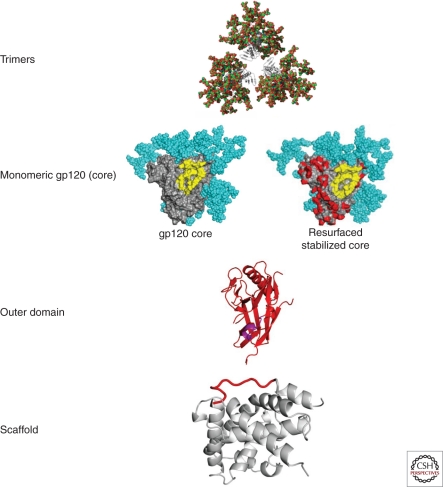
The development of a highly effective AIDS vaccine will likely depend on success in designing immunogens that elicit broadly neutralizing antibodies to naturally circulating strains of HIV-1. Although the antibodies induced after natural infection with HIV-1 are often directed to strain-specific or nonneutralizing determinants, it is now evident that 10%-25% of HIV-infected individuals generate neutralizing antibody responses of considerable breadth. In the past, only four broadly neutralizing monoclonal antibodies had been defined, but more than a dozen monoclonal antibodies of substantial breadth have more recently been isolated. An understanding of their recognition sites, the structural basis of their interaction with the HIV Env, and their development pathways provides new opportunities to design vaccine candidates that will elicit broadly protective antibodies against this virus.
Figures
Similar articles
-
Conformational Epitope-Specific Broadly Neutralizing Plasma Antibodies Obtained from an HIV-1 Clade C-Infected Elite Neutralizer Mediate Autologous Virus Escape through Mutations in the V1 Loop.J Virol. 2016 Jan 13;90(7):3446-57. doi: 10.1128/JVI.03090-15. J Virol. 2016. PMID: 26763999 Free PMC article.
-
Rapid Induction of Multifunctional Antibodies in Rabbits and Macaques by Clade C HIV-1 CAP257 Envelopes Circulating During Epitope-Specific Neutralization Breadth Development.Front Immunol. 2020 Jun 2;11:984. doi: 10.3389/fimmu.2020.00984. eCollection 2020. Front Immunol. 2020. PMID: 32582155 Free PMC article.
-
The Neutralizing Antibody Response to the HIV-1 Env Protein.Curr HIV Res. 2018;16(1):21-28. doi: 10.2174/1570162X15666171124122044. Curr HIV Res. 2018. PMID: 29173180 Free PMC article. Review.
-
Broadly neutralizing antibodies: What is needed to move from a rare event in HIV-1 infection to vaccine efficacy?Retrovirology. 2018 Jul 28;15(1):52. doi: 10.1186/s12977-018-0433-2. Retrovirology. 2018. PMID: 30055627 Free PMC article. Review.
-
Neutralizing antibodies and control of HIV: moves and countermoves.Curr HIV/AIDS Rep. 2012 Mar;9(1):64-72. doi: 10.1007/s11904-011-0105-5. Curr HIV/AIDS Rep. 2012. PMID: 22203469 Review.
Cited by
-
Comparison of viral Env proteins from acute and chronic infections with subtype C human immunodeficiency virus type 1 identifies differences in glycosylation and CCR5 utilization and suggests a new strategy for immunogen design.J Virol. 2013 Jul;87(13):7218-33. doi: 10.1128/JVI.03577-12. Epub 2013 Apr 24. J Virol. 2013. PMID: 23616655 Free PMC article.
-
Safety and immunogenicity of DNA prime and modified vaccinia ankara virus-HIV subtype C vaccine boost in healthy adults.Clin Vaccine Immunol. 2013 Mar;20(3):397-408. doi: 10.1128/CVI.00637-12. Epub 2013 Jan 23. Clin Vaccine Immunol. 2013. PMID: 23345581 Free PMC article. Clinical Trial.
-
Frequencies of Circulating Th1-Biased T Follicular Helper Cells in Acute HIV-1 Infection Correlate with the Development of HIV-Specific Antibody Responses and Lower Set Point Viral Load.J Virol. 2018 Jul 17;92(15):e00659-18. doi: 10.1128/JVI.00659-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29793949 Free PMC article.
-
COVID-19 Vaccines Currently under Preclinical and Clinical Studies, and Associated Antiviral Immune Response.Vaccines (Basel). 2020 Nov 3;8(4):649. doi: 10.3390/vaccines8040649. Vaccines (Basel). 2020. PMID: 33153096 Free PMC article. Review.
-
A Blueprint for HIV Vaccine Discovery.Cell Host Microbe. 2012 Oct 18;12(4):396-407. doi: 10.1016/j.chom.2012.09.008. Cell Host Microbe. 2012. PMID: 23084910 Free PMC article. Review.
References
-
- Alam SM, McAdams M, Boren D, Rak M, Scearce RM, Gao F, Camacho ZT, Gewirth D, Kelsoe G, Chen P, et al. 2007. The role of antibody polyspecificity and lipid reactivity in binding of broadly neutralizing anti-HIV-1 envelope human monoclonal antibodies 2F5 and 4E10 to glycoprotein 41 membrane proximal envelope epitopes. J Immunol 178: 4424–4435 - PMC - PubMed
-
- Albert J, Abrahamsson B, Nagy K, Aurelius E, Gaines H, Nystrom G, Fenyo EM 1990. Rapid development of isolate-specific neutralizing antibodies after primary HIV-1 infection and consequent emergence of virus variants which resist neutralization by autologous sera. AIDS 4: 107–112 - PubMed
-
- Arthos J, Cicala C, Martinelli E, Macleod K, Van Ryk D, Wei D, Xiao Z, Veenstra TD, Conrad TP, Lempicki RA, et al. 2008. HIV-1 envelope protein binds to and signals through integrin α4β7, the gut mucosal homing receptor for peripheral T cells. Nat Immunol 9: 301–309 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous