

Peroxide-dependent sulfenylation of the EGFR catalytic site enhances kinase activity
- PMID: 22158416
- PMCID: PMC3528018
- DOI: 10.1038/nchembio.736
Peroxide-dependent sulfenylation of the EGFR catalytic site enhances kinase activity
Abstract
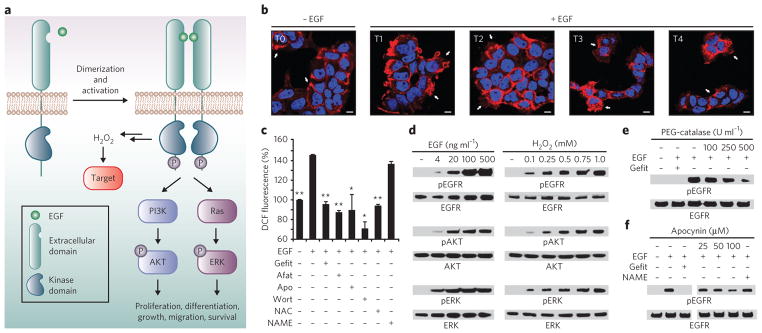
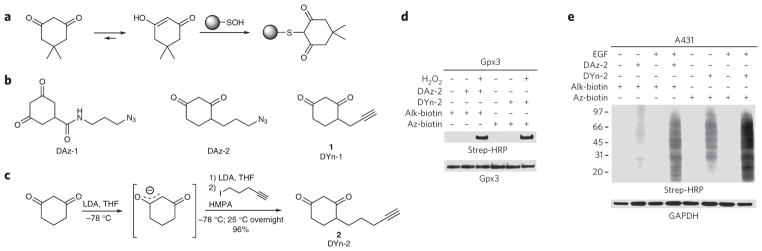
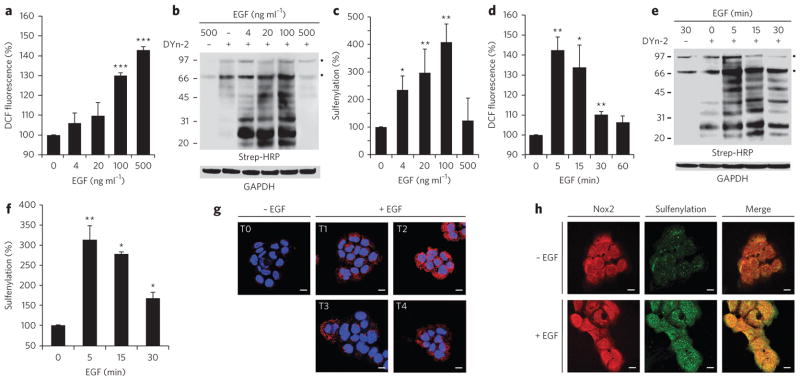
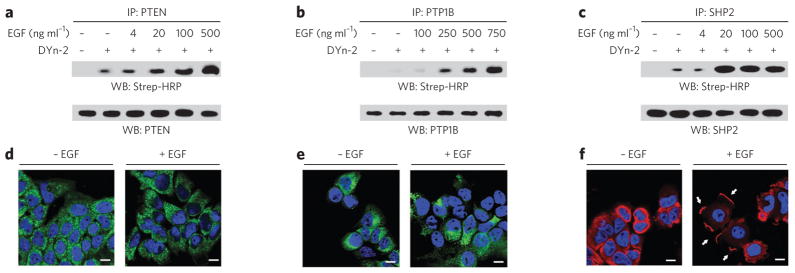
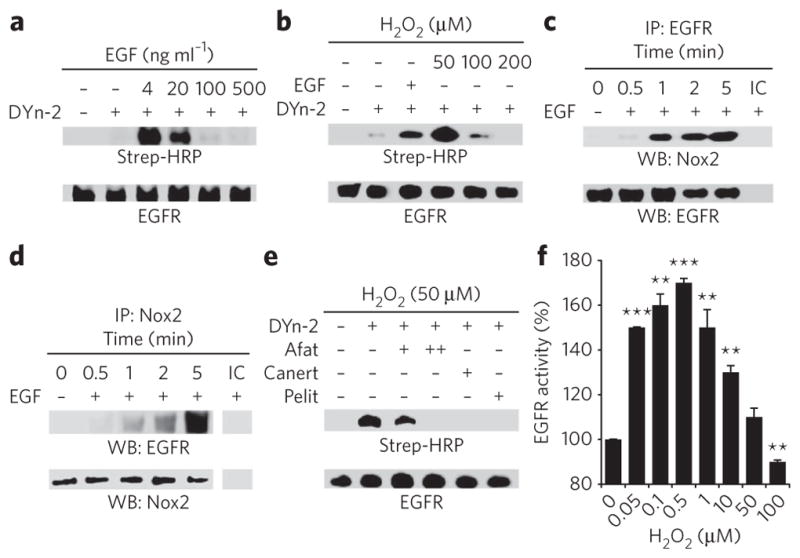
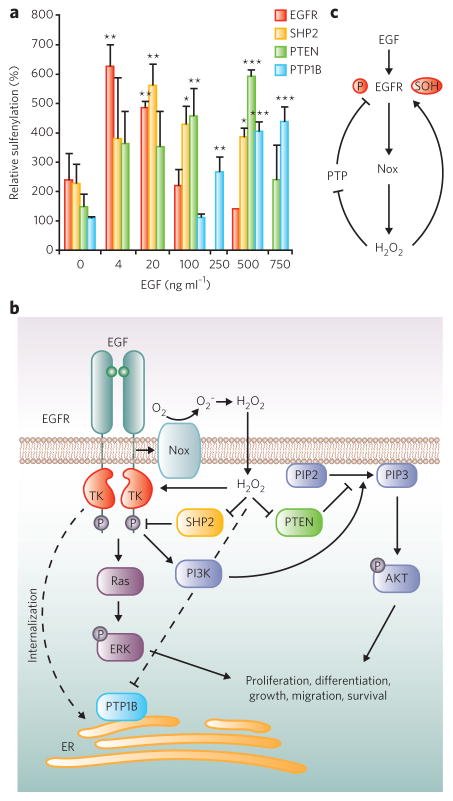
Protein sulfenylation is a post-translational modification of emerging importance in higher eukaryotes. However, investigation of its diverse roles remains challenging, particularly within a native cellular environment. Herein we report the development and application of DYn-2, a new chemoselective probe for detecting sulfenylated proteins in human cells. These studies show that epidermal growth factor receptor-mediated signaling results in H(2)O(2) production and oxidation of downstream proteins. In addition, we demonstrate that DYn-2 has the ability to detect differences in sulfenylation rates within the cell, which are associated with differences in target protein localization. We also show that the direct modification of epidermal growth factor receptor by H(2)O(2) at a critical active site cysteine (Cys797) enhances its tyrosine kinase activity. Collectively, our findings reveal sulfenylation as a global signaling mechanism that is akin to phosphorylation and has regulatory implications for other receptor tyrosine kinases and irreversible inhibitors that target oxidant-sensitive cysteines in proteins.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Mining for protein S-sulfenylation in Arabidopsis uncovers redox-sensitive sites.Proc Natl Acad Sci U S A. 2019 Oct 15;116(42):21256-21261. doi: 10.1073/pnas.1906768116. Epub 2019 Oct 2. Proc Natl Acad Sci U S A. 2019. PMID: 31578252 Free PMC article.
-
Redox regulation of epidermal growth factor receptor signaling through cysteine oxidation.Biochemistry. 2012 Dec 18;51(50):9954-65. doi: 10.1021/bi301441e. Epub 2012 Dec 5. Biochemistry. 2012. PMID: 23186290 Free PMC article. Review.
-
The NADPH Oxidases DUOX1 and NOX2 Play Distinct Roles in Redox Regulation of Epidermal Growth Factor Receptor Signaling.J Biol Chem. 2016 Oct 28;291(44):23282-23293. doi: 10.1074/jbc.M116.749028. Epub 2016 Sep 20. J Biol Chem. 2016. PMID: 27650496 Free PMC article.
-
Hydrogen peroxide generated during cellular insulin stimulation is integral to activation of the distal insulin signaling cascade in 3T3-L1 adipocytes.J Biol Chem. 2001 Dec 28;276(52):48662-9. doi: 10.1074/jbc.M105061200. Epub 2001 Oct 11. J Biol Chem. 2001. PMID: 11598110
-
Chemical biology approaches to study protein cysteine sulfenylation.Biopolymers. 2014 Feb;101(2):165-72. doi: 10.1002/bip.22255. Biopolymers. 2014. PMID: 23576224 Free PMC article. Review.
Cited by
-
Oxidation-Dependent Activation of Src Kinase Mediates Epithelial IL-33 Production and Signaling during Acute Airway Allergen Challenge.J Immunol. 2021 Jun 15;206(12):2989-2999. doi: 10.4049/jimmunol.2000995. Epub 2021 Jun 4. J Immunol. 2021. PMID: 34088769 Free PMC article.
-
Bifunctional electrophiles cross-link thioredoxins with redox relay partners in cells.Chem Res Toxicol. 2013 Mar 18;26(3):490-7. doi: 10.1021/tx4000123. Epub 2013 Mar 4. Chem Res Toxicol. 2013. PMID: 23414292 Free PMC article.
-
Sulfur signaling pathway in cardiovascular disease.Front Pharmacol. 2023 Nov 24;14:1303465. doi: 10.3389/fphar.2023.1303465. eCollection 2023. Front Pharmacol. 2023. PMID: 38074127 Free PMC article. Review.
-
Pro-Oxidant Auranofin and Glutathione-Depleting Combination Unveils Synergistic Lethality in Glioblastoma Cells with Aberrant Epidermal Growth Factor Receptor Expression.Cancers (Basel). 2024 Jun 25;16(13):2319. doi: 10.3390/cancers16132319. Cancers (Basel). 2024. PMID: 39001381 Free PMC article.
-
Chemoselective ligation of sulfinic acids with aryl-nitroso compounds.Angew Chem Int Ed Engl. 2012 Jun 25;51(26):6502-5. doi: 10.1002/anie.201201812. Epub 2012 May 29. Angew Chem Int Ed Engl. 2012. PMID: 22644884 Free PMC article.
References
-
- Miller EW, Tulyathan O, Isacoff EY, Chang CJ. Molecular imaging of hydrogen peroxide produced for cell signaling. Nat Chem Biol. 2007;3:263–267. erratum 3 349 2007. - PubMed
-
- Woo HA, et al. Inactivation of peroxiredoxin I by phosphorylation allows localized H2O2 accumulation for cell signaling. Cell. 2010;140:517–528. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
