

Extramedullary hematopoiesis generates Ly-6C(high) monocytes that infiltrate atherosclerotic lesions
- PMID: 22144566
- PMCID: PMC3263762
- DOI: 10.1161/CIRCULATIONAHA.111.061986
Extramedullary hematopoiesis generates Ly-6C(high) monocytes that infiltrate atherosclerotic lesions
Abstract
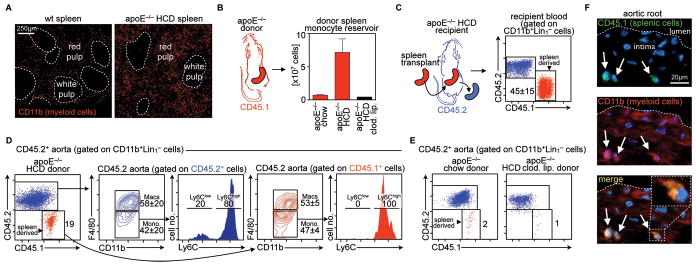
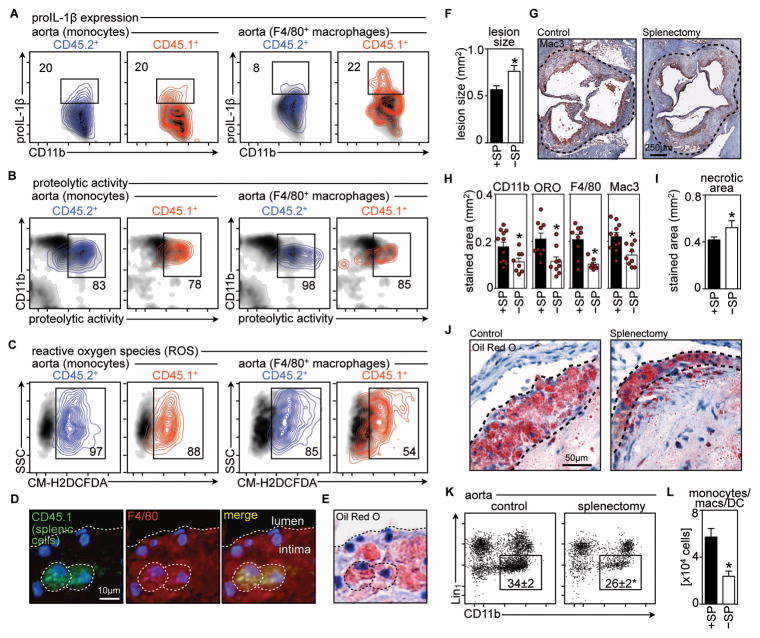
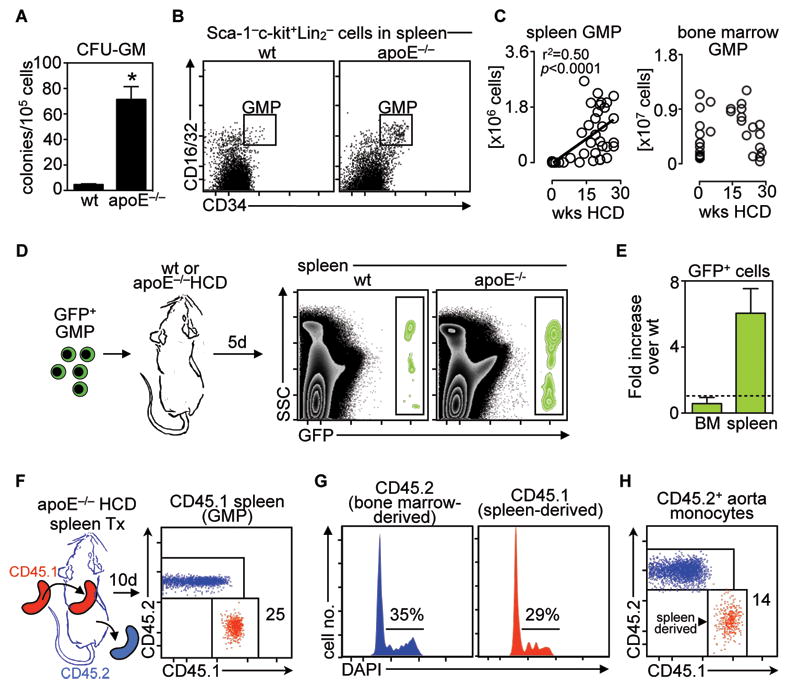
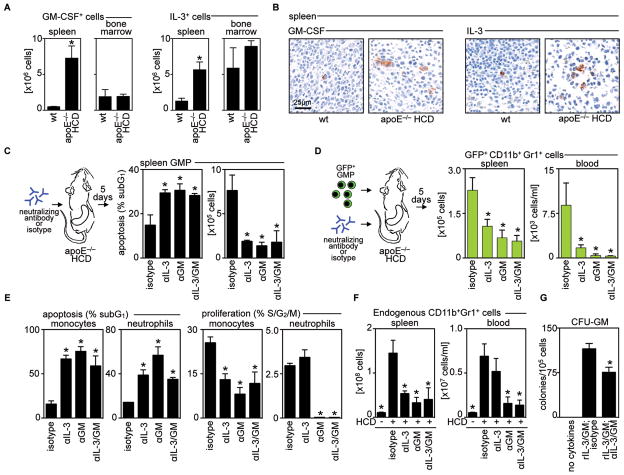
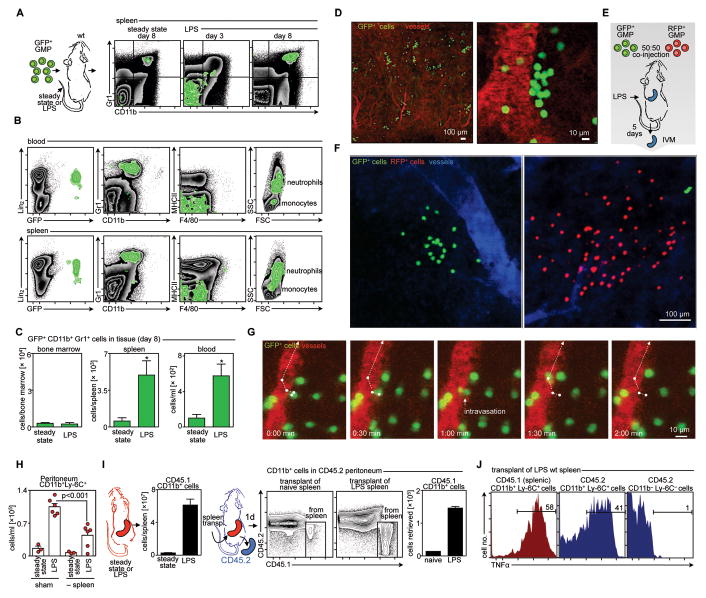
Background: Atherosclerotic lesions are believed to grow via the recruitment of bone marrow-derived monocytes. Among the known murine monocyte subsets, Ly-6C(high) monocytes are inflammatory, accumulate in lesions preferentially, and differentiate. Here, we hypothesized that the bone marrow outsources the production of Ly-6C(high) monocytes during atherosclerosis.
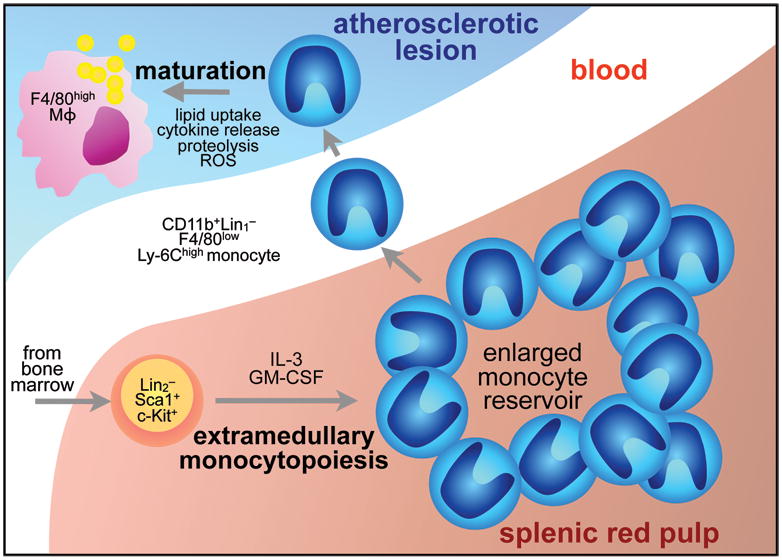
Methods and results: Using murine models of atherosclerosis and fate-mapping approaches, we show that hematopoietic stem and progenitor cells progressively relocate from the bone marrow to the splenic red pulp, where they encounter granulocyte macrophage colony-stimulating factor and interleukin-3, clonally expand, and differentiate to Ly-6C(high) monocytes. Monocytes born in such extramedullary niches intravasate, circulate, and accumulate abundantly in atheromata. On lesional infiltration, Ly-6C(high) monocytes secrete inflammatory cytokines, reactive oxygen species, and proteases. Eventually, they ingest lipids and become foam cells.
Conclusions: Our findings indicate that extramedullary sites supplement the hematopoietic function of the bone marrow by producing circulating inflammatory cells that infiltrate atherosclerotic lesions.
© 2011 American Heart Association, Inc.
Conflict of interest statement
Figures

Similar articles
-
P-selectin glycoprotein ligand-1 is highly expressed on Ly-6Chi monocytes and a major determinant for Ly-6Chi monocyte recruitment to sites of atherosclerosis in mice.Circulation. 2008 Jun 24;117(25):3227-37. doi: 10.1161/CIRCULATIONAHA.108.771048. Epub 2008 Jun 2. Circulation. 2008. PMID: 18519846 Free PMC article.
-
Accumulation of CD11b+ lung dendritic cells in response to fungal infection results from the CCR2-mediated recruitment and differentiation of Ly-6Chigh monocytes.J Immunol. 2009 Dec 15;183(12):8044-53. doi: 10.4049/jimmunol.0902823. J Immunol. 2009. PMID: 19933856 Free PMC article.
-
Impaired infarct healing in atherosclerotic mice with Ly-6C(hi) monocytosis.J Am Coll Cardiol. 2010 Apr 13;55(15):1629-38. doi: 10.1016/j.jacc.2009.08.089. J Am Coll Cardiol. 2010. PMID: 20378083 Free PMC article.
-
Monocyte and macrophage dynamics during atherogenesis.Arterioscler Thromb Vasc Biol. 2011 Jul;31(7):1506-16. doi: 10.1161/ATVBAHA.110.221127. Arterioscler Thromb Vasc Biol. 2011. PMID: 21677293 Free PMC article. Review.
-
Monocyte fate in atherosclerosis.Arterioscler Thromb Vasc Biol. 2015 Feb;35(2):272-9. doi: 10.1161/ATVBAHA.114.303565. Epub 2014 Dec 23. Arterioscler Thromb Vasc Biol. 2015. PMID: 25538208 Review.
Cited by
-
Monocyte-directed RNAi targeting CCR2 improves infarct healing in atherosclerosis-prone mice.Circulation. 2013 May 21;127(20):2038-46. doi: 10.1161/CIRCULATIONAHA.112.000116. Epub 2013 Apr 24. Circulation. 2013. PMID: 23616627 Free PMC article.
-
Leukocyte behavior in atherosclerosis, myocardial infarction, and heart failure.Science. 2013 Jan 11;339(6116):161-6. doi: 10.1126/science.1230719. Science. 2013. PMID: 23307733 Free PMC article. Review.
-
Monocyte and macrophage heterogeneity in the heart.Circ Res. 2013 Jun 7;112(12):1624-33. doi: 10.1161/CIRCRESAHA.113.300890. Circ Res. 2013. PMID: 23743228 Free PMC article. Review.
-
Glucocorticoid-induced tumour necrosis factor receptor family-related protein (GITR) drives atherosclerosis in mice and is associated with an unstable plaque phenotype and cerebrovascular events in humans.Eur Heart J. 2020 Aug 14;41(31):2938-2948. doi: 10.1093/eurheartj/ehaa484. Eur Heart J. 2020. PMID: 32728688 Free PMC article.
-
Imaging systemic inflammatory networks in ischemic heart disease.J Am Coll Cardiol. 2015 Apr 21;65(15):1583-91. doi: 10.1016/j.jacc.2015.02.034. J Am Coll Cardiol. 2015. PMID: 25881940 Free PMC article. Review.
References
-
- Medzhitov R. Origin and physiological roles of inflammation. Nature. 2008;454:428–435. - PubMed
-
- Auffray C, Sieweke MH, Geissmann F. Blood monocytes: development, heterogeneity, and relationship with dendritic cells. Annu Rev Immunol. 2009;27:669–692. - PubMed
-
- Nathan C. Neutrophils and immunity: challenges and opportunities. Nat Rev Immunol. 2006;6:173–182. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL095612/HL/NHLBI NIH HHS/United States
- U24 CA092782/CA/NCI NIH HHS/United States
- U01 HL080731/HL/NHLBI NIH HHS/United States
- R01 AI084880/AI/NIAID NIH HHS/United States
- R01 HL095612-02/HL/NHLBI NIH HHS/United States
- 1R01 HL095612/HL/NHLBI NIH HHS/United States
- P01-A154904/PHS HHS/United States
- U24 CA092782-07/CA/NCI NIH HHS/United States
- P50 CA86355/CA/NCI NIH HHS/United States
- R01 HL069948-01A1/HL/NHLBI NIH HHS/United States
- P50 CA086355-01/CA/NCI NIH HHS/United States
- P01 AI054904/AI/NIAID NIH HHS/United States
- P50 CA086355/CA/NCI NIH HHS/United States
- 2R01 HL069948/HL/NHLBI NIH HHS/United States
- R01 HL069948/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
