

Calcium- and integrin-binding protein 1 regulates megakaryocyte ploidy, adhesion, and migration
- PMID: 22128142
- PMCID: PMC3265206
- DOI: 10.1182/blood-2011-04-346098
Calcium- and integrin-binding protein 1 regulates megakaryocyte ploidy, adhesion, and migration
Abstract
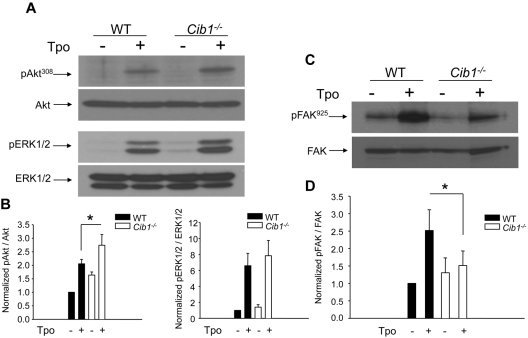
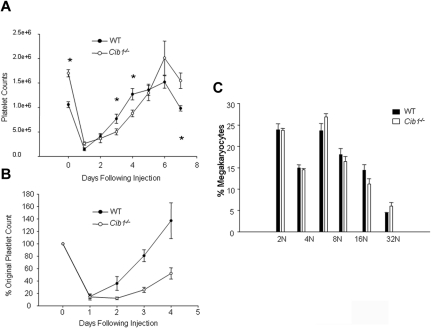
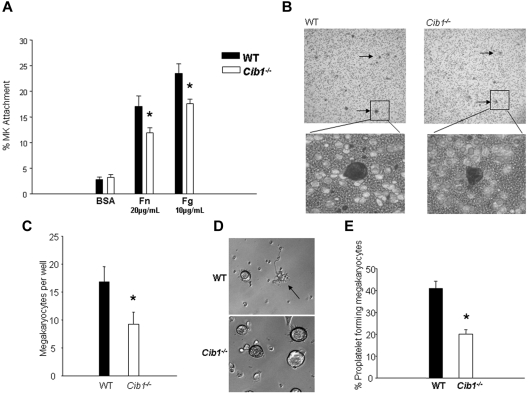
Megakaryocytes are large, polyploid cells that produce platelets. We have previously reported that calcium- and integrin-binding protein 1 (CIB1) regulates endomitosis in Dami cells. To further characterize the role of CIB1 in megakaryopoiesis, we used a Cib1(-/-) mouse model. Cib1(-/-) mice have more platelets and BM megakaryocytes than wild-type (WT) controls (P < .05). Furthermore, subsequent analysis of megakaryocyte-CFU production revealed an increase with Cib1 deletion compared with WT (P < .05). In addition, BM from Cib1(-/-) mice, cultured with thrombopoietin (TPO) for 24 hours, produced more highly polyploid megakaryocytes than WT BM (P < .05). Subsequent analysis of TPO signaling revealed enhanced Akt and ERK1/2 phosphorylation, whereas FAK(Y925) phosphorylation was reduced in Cib1(-/-) megakaryocytes treated with TPO. Conversely, platelet recovery in Cib1(-/-) mice after platelet depletion was attenuated compared with WT (P < .05). This could be the result of impaired adhesion and migration, as adhesion to fibrinogen and fibronectin and migration toward an SDF-1α gradient were reduced in Cib1(-/-) megakaryocytes compared with WT (P < .05). In addition, Cib1(-/-) megakaryocytes formed fewer proplatelets compared with WT (P < .05), when plated on fibrinogen. These data suggest that CIB1 plays a dual role in megakaryopoiesis, initially by negatively regulating TPO signaling and later by augmenting proplatelet production.
Figures
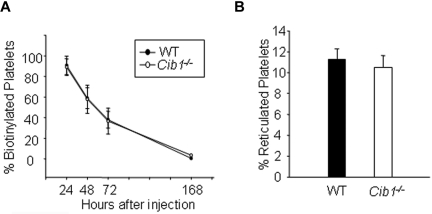
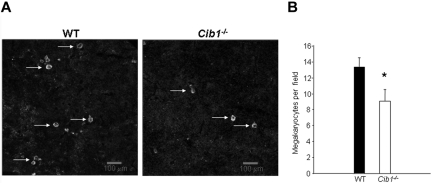
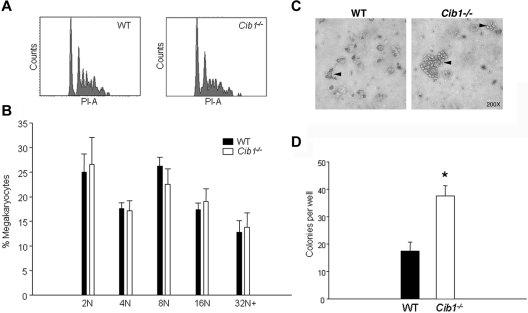
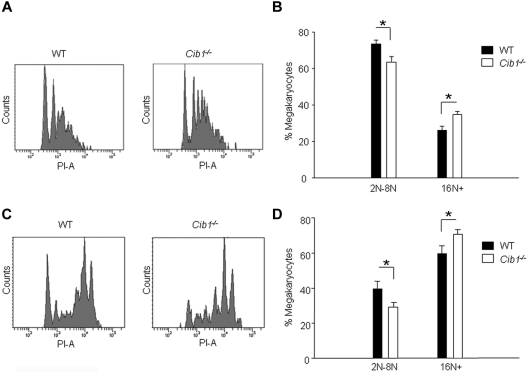
Similar articles
-
Roles of focal adhesion kinase (FAK) in megakaryopoiesis and platelet function: studies using a megakaryocyte lineage specific FAK knockout.Blood. 2008 Jan 15;111(2):596-604. doi: 10.1182/blood-2007-05-089680. Epub 2007 Oct 9. Blood. 2008. PMID: 17925492 Free PMC article.
-
Calcium- and integrin-binding protein 1 regulates endomitosis and its interaction with Polo-like kinase 3 is enhanced in endomitotic Dami cells.PLoS One. 2011 Jan 14;6(1):e14513. doi: 10.1371/journal.pone.0014513. PLoS One. 2011. PMID: 21264284 Free PMC article.
-
Thrombopoietin-induced activation of the mitogen-activated protein kinase (MAPK) pathway in normal megakaryocytes: role in endomitosis.Blood. 1999 Aug 15;94(4):1273-82. Blood. 1999. PMID: 10438715
-
Milestones in understanding platelet production: a historical overview.Br J Haematol. 2014 Apr;165(2):248-58. doi: 10.1111/bjh.12781. Epub 2014 Feb 14. Br J Haematol. 2014. PMID: 24528208 Review.
-
Megakaryocytes and beyond: the birth of platelets.J Thromb Haemost. 2003 Jun;1(6):1174-82. doi: 10.1046/j.1538-7836.2003.00290.x. J Thromb Haemost. 2003. PMID: 12871316 Review.
Cited by
-
Discovery and Development of Cyclic Peptide Inhibitors of CIB1.ACS Med Chem Lett. 2021 Oct 27;12(11):1832-1839. doi: 10.1021/acsmedchemlett.1c00438. eCollection 2021 Nov 11. ACS Med Chem Lett. 2021. PMID: 34795874 Free PMC article.
-
iTRAQ based quantitative proteomics approach validated the role of calcyclin binding protein (CacyBP) in promoting colorectal cancer metastasis.Mol Cell Proteomics. 2013 Jul;12(7):1865-80. doi: 10.1074/mcp.M112.023085. Epub 2013 Mar 29. Mol Cell Proteomics. 2013. PMID: 23543800 Free PMC article.
-
Calcium- and integrin-binding protein-1 is down-regulated in the sperm of patients with oligoasthenozoospermia : CIB1 expression in patients with oligoasthenozoospermia.J Assist Reprod Genet. 2014 May;31(5):541-7. doi: 10.1007/s10815-014-0177-4. Epub 2014 Jan 26. J Assist Reprod Genet. 2014. PMID: 24464679 Free PMC article.
-
Ask1 regulates murine platelet granule secretion, thromboxane A2 generation, and thrombus formation.Blood. 2017 Mar 2;129(9):1197-1209. doi: 10.1182/blood-2016-07-729780. Epub 2016 Dec 27. Blood. 2017. PMID: 28028021 Free PMC article.
-
CIB1 synergizes with EphrinA2 to regulate Kaposi's sarcoma-associated herpesvirus macropinocytic entry in human microvascular dermal endothelial cells.PLoS Pathog. 2014 Feb 13;10(2):e1003941. doi: 10.1371/journal.ppat.1003941. eCollection 2014 Feb. PLoS Pathog. 2014. PMID: 24550731 Free PMC article.
References
-
- Junt T, Schulze H, Chen Z, et al. Dynamic visualization of thrombopoiesis within bone marrow. Science. 2007;317(5845):1767–1770. - PubMed
-
- de Sauvage FJ, Hass PE, Spencer SD, et al. Stimulation of megakaryocytopoiesis and thrombopoiesis by the c-Mpl ligand. Nature. 1994;369(6481):533–538. - PubMed
-
- Nakao T, Geddis AE, Fox NE, Kaushansky K. PI3K/Akt/FOXO3a pathway contributes to thrombopoietin-induced proliferation of primary megakaryocytes in vitro and in vivo via modulation of p27(Kip1). Cell Cycle. 2008;7(2):257–266. - PubMed
-
- Rojnuckarin P, Drachman JG, Kaushansky K. Thrombopoietin-induced activation of the mitogen-activated protein kinase (MAPK) pathway in normal megakaryocytes: role in endomitosis. Blood. 1999;94(4):1273–1282. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
