

rRNA pseudouridylation defects affect ribosomal ligand binding and translational fidelity from yeast to human cells
- PMID: 22099312
- PMCID: PMC3222873
- DOI: 10.1016/j.molcel.2011.09.017
rRNA pseudouridylation defects affect ribosomal ligand binding and translational fidelity from yeast to human cells
Abstract
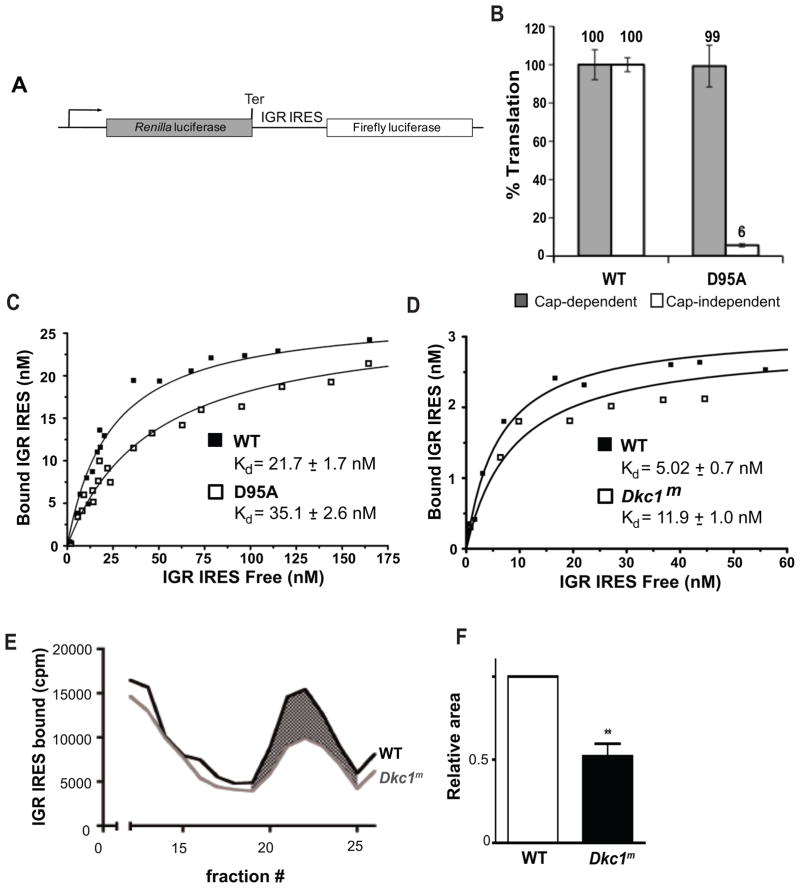
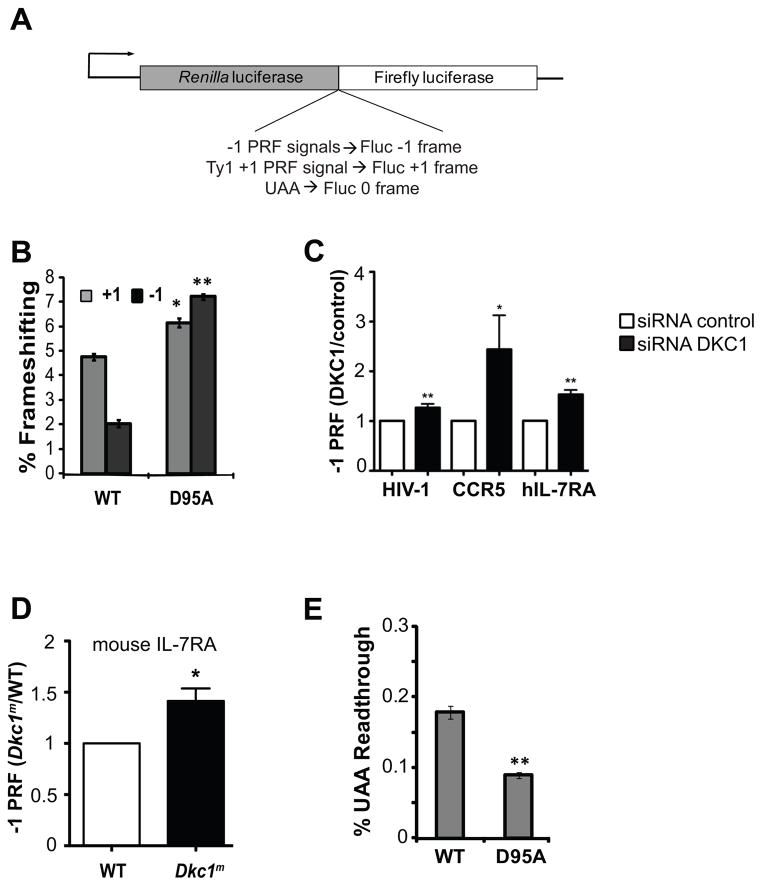
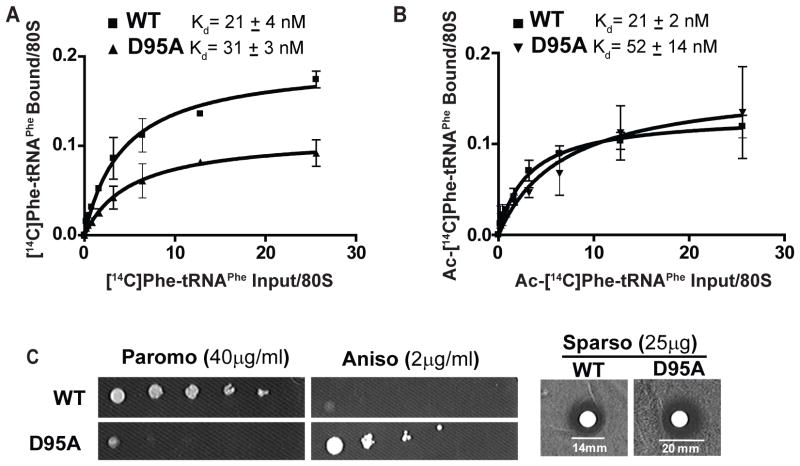
How pseudouridylation (Ψ), the most common and evolutionarily conserved modification of rRNA, regulates ribosome activity is poorly understood. Medically, Ψ is important because the rRNA Ψ synthase, DKC1, is mutated in X-linked dyskeratosis congenita (X-DC) and Hoyeraal-Hreidarsson (HH) syndrome. Here, we characterize ribosomes isolated from a yeast strain in which Cbf5p, the yeast homolog of DKC1, is catalytically impaired through a D95A mutation (cbf5-D95A). Ribosomes from cbf5-D95A cells display decreased affinities for tRNA binding to the A and P sites as well as the cricket paralysis virus internal ribosome entry site (IRES), which interacts with both the P and the E sites of the ribosome. This biochemical impairment in ribosome activity manifests as decreased translational fidelity and IRES-dependent translational initiation, which are also evident in mouse and human cells deficient for DKC1 activity. These findings uncover specific roles for Ψ modification in ribosome-ligand interactions that are conserved in yeast, mouse, and humans.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
RNA-guided isomerization of uridine to pseudouridine--pseudouridylation.RNA Biol. 2014;11(12):1483-94. doi: 10.4161/15476286.2014.972855. RNA Biol. 2014. PMID: 25590339 Free PMC article. Review.
-
Point mutations in yeast CBF5 can abolish in vivo pseudouridylation of rRNA.Mol Cell Biol. 1999 Nov;19(11):7461-72. doi: 10.1128/MCB.19.11.7461. Mol Cell Biol. 1999. PMID: 10523634 Free PMC article.
-
Severity of X-linked dyskeratosis congenita (DKCX) cellular defects is not directly related to dyskerin (DKC1) activity in ribosomal RNA biogenesis or mRNA translation.Hum Mutat. 2013 Dec;34(12):1698-707. doi: 10.1002/humu.22447. Epub 2013 Oct 21. Hum Mutat. 2013. PMID: 24115260
-
The yeast nucleolar protein Cbf5p is involved in rRNA biosynthesis and interacts genetically with the RNA polymerase I transcription factor RRN3.Mol Cell Biol. 1997 Oct;17(10):6175-83. doi: 10.1128/MCB.17.10.6175. Mol Cell Biol. 1997. PMID: 9315678 Free PMC article.
-
Turning Uridines around: Role of rRNA Pseudouridylation in Ribosome Biogenesis and Ribosomal Function.Biomolecules. 2018 Jun 5;8(2):38. doi: 10.3390/biom8020038. Biomolecules. 2018. PMID: 29874862 Free PMC article. Review.
Cited by
-
A reconstituted cell-free assay for the evaluation of the intrinsic activity of purified human ribosomes.Nat Protoc. 2016 Jul;11(7):1309-25. doi: 10.1038/nprot.2016.072. Epub 2016 Jun 23. Nat Protoc. 2016. PMID: 27336708
-
PSI-MOUSE: Predicting Mouse Pseudouridine Sites From Sequence and Genome-Derived Features.Evol Bioinform Online. 2020 Jun 9;16:1176934320925752. doi: 10.1177/1176934320925752. eCollection 2020. Evol Bioinform Online. 2020. PMID: 32565674 Free PMC article.
-
Multiple Parallel Pathways of Translation Initiation on the CrPV IRES.Mol Cell. 2016 Apr 7;62(1):92-103. doi: 10.1016/j.molcel.2016.03.020. Mol Cell. 2016. PMID: 27058789 Free PMC article.
-
RNA-guided isomerization of uridine to pseudouridine--pseudouridylation.RNA Biol. 2014;11(12):1483-94. doi: 10.4161/15476286.2014.972855. RNA Biol. 2014. PMID: 25590339 Free PMC article. Review.
-
RNA pseudouridylation: new insights into an old modification.Trends Biochem Sci. 2013 Apr;38(4):210-8. doi: 10.1016/j.tibs.2013.01.002. Epub 2013 Feb 4. Trends Biochem Sci. 2013. PMID: 23391857 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- R01GM084547/GM/NIGMS NIH HHS/United States
- R01 GM058859/GM/NIGMS NIH HHS/United States
- 3R01GM084547-01A1S1/GM/NIGMS NIH HHS/United States
- R01 GM084547-01A1S1/GM/NIGMS NIH HHS/United States
- R01 HL085572-05/HL/NHLBI NIH HHS/United States
- R01 GM058859-10A1S1/GM/NIGMS NIH HHS/United States
- R01 GM058859-13/GM/NIGMS NIH HHS/United States
- R01HL085572/HL/NHLBI NIH HHS/United States
- 5R01GM058859/GM/NIGMS NIH HHS/United States
- R01 DK098057/DK/NIDDK NIH HHS/United States
- R01 HL085572-04/HL/NHLBI NIH HHS/United States
- R01 GM084547/GM/NIGMS NIH HHS/United States
- R01 GM084547-03/GM/NIGMS NIH HHS/United States
- 3R01GM058859-10A1S1/GM/NIGMS NIH HHS/United States
- R01 HL085572/HL/NHLBI NIH HHS/United States
- R01 GM084547-02/GM/NIGMS NIH HHS/United States
- R01 GM058859-12/GM/NIGMS NIH HHS/United States
- 3R01HL085572-05S1/HL/NHLBI NIH HHS/United States
- R01 GM084547-04/GM/NIGMS NIH HHS/United States
- R01 HL085572-05S1/HL/NHLBI NIH HHS/United States
- R01 GM058859-10A1/GM/NIGMS NIH HHS/United States
- R01 GM058859-11/GM/NIGMS NIH HHS/United States
- R01 HL085572-03/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
