

CBP mediates NF-κB-dependent histone acetylation and estrogen receptor recruitment to an estrogen response element in the BIRC3 promoter
- PMID: 22083956
- PMCID: PMC3255764
- DOI: 10.1128/MCB.05869-11
CBP mediates NF-κB-dependent histone acetylation and estrogen receptor recruitment to an estrogen response element in the BIRC3 promoter
Abstract
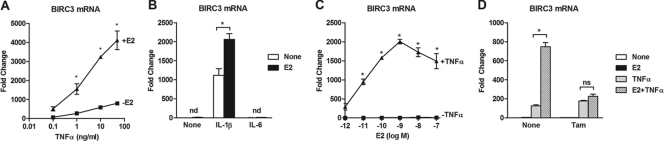
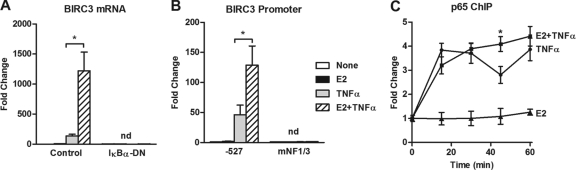
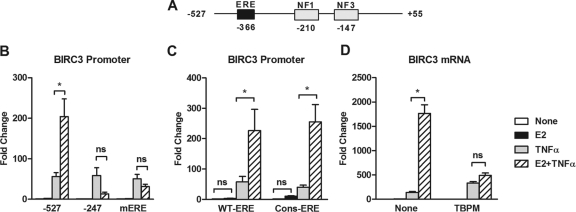
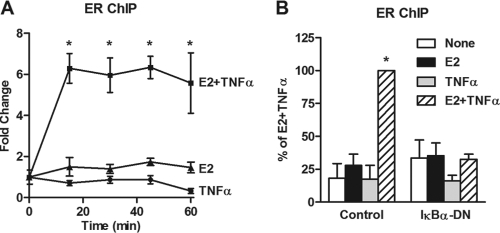
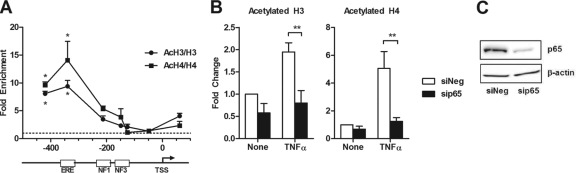
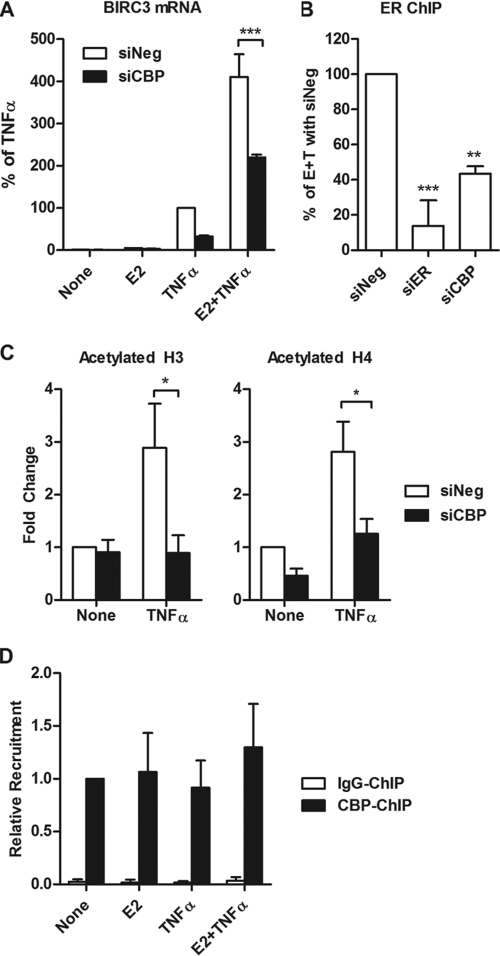
Estrogen receptor (ER) and NF-κB are transcription factors with profound effects on breast cancer cell proliferation and survival. While many studies demonstrate that ER and NF-κB can repress each other, we previously identified a gene signature that is synergistically upregulated by these two factors in more aggressive luminal B breast tumors. Herein, we examine a novel mechanism of cross talk between ER and NF-κB that results in the upregulation of the antiapoptotic gene BIRC3 (also known as cIAP2). We demonstrate that NF-κB, acting through two response elements, is required for ER recruitment to an adjacent estrogen response element (ERE) in the BIRC3 promoter. This effect is accompanied by a major increase in NF-κB-dependent histone acetylation around the ERE. Interestingly, CBP, a histone acetyltransferase previously implicated in repressive interactions between ER and NF-κB, plays a permissive role by promoting histone acetylation and ER recruitment, as well as enhanced expression of BIRC3. These findings suggest a new gene regulatory mechanism by which inflammation and NF-κB activation can influence ER recruitment to inherently inactive ER binding sites. This fine-tuning mechanism may explain how two factors that generally repress each other's activity may work together on certain genes to promote breast cancer cell survival and tumor progression.
Figures
Similar articles
-
NFκB affects estrogen receptor expression and activity in breast cancer through multiple mechanisms.Mol Cell Endocrinol. 2015 Dec 15;418 Pt 3(0 3):235-9. doi: 10.1016/j.mce.2014.09.013. Epub 2014 Oct 18. Mol Cell Endocrinol. 2015. PMID: 25450861 Free PMC article. Review.
-
Positive cross-talk between estrogen receptor and NF-kappaB in breast cancer.Cancer Res. 2009 Dec 1;69(23):8918-25. doi: 10.1158/0008-5472.CAN-09-2608. Epub 2009 Nov 17. Cancer Res. 2009. PMID: 19920189 Free PMC article.
-
Proinflammatory cytokines enhance estrogen-dependent expression of the multidrug transporter gene ABCG2 through estrogen receptor and NF{kappa}B cooperativity at adjacent response elements.J Biol Chem. 2010 Oct 8;285(41):31100-6. doi: 10.1074/jbc.M110.155309. Epub 2010 Aug 12. J Biol Chem. 2010. PMID: 20705611 Free PMC article.
-
Up-Regulation of Glioma-Associated Oncogene Homolog 1 Expression by Serum Starvation Promotes Cell Survival in ER-Positive Breast Cancer Cells.Cell Physiol Biochem. 2015;36(5):1862-76. doi: 10.1159/000430156. Cell Physiol Biochem. 2015. PMID: 26182949
-
Linkage between EGFR family receptors and nuclear factor kappaB (NF-kappaB) signaling in breast cancer.J Cell Physiol. 2006 Dec;209(3):645-52. doi: 10.1002/jcp.20785. J Cell Physiol. 2006. PMID: 17001676 Review.
Cited by
-
Novel ceRNA network construction associated with programmed cell death in acute rejection of heart allograft in mice.Front Immunol. 2023 Sep 11;14:1184409. doi: 10.3389/fimmu.2023.1184409. eCollection 2023. Front Immunol. 2023. PMID: 37753085 Free PMC article.
-
NFκB affects estrogen receptor expression and activity in breast cancer through multiple mechanisms.Mol Cell Endocrinol. 2015 Dec 15;418 Pt 3(0 3):235-9. doi: 10.1016/j.mce.2014.09.013. Epub 2014 Oct 18. Mol Cell Endocrinol. 2015. PMID: 25450861 Free PMC article. Review.
-
Targeting Histone Modifications in Breast Cancer: A Precise Weapon on the Way.Front Cell Dev Biol. 2021 Sep 14;9:736935. doi: 10.3389/fcell.2021.736935. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34595180 Free PMC article. Review.
-
Structural and Molecular Mechanisms of Cytokine-Mediated Endocrine Resistance in Human Breast Cancer Cells.Mol Cell. 2017 Mar 16;65(6):1122-1135.e5. doi: 10.1016/j.molcel.2017.02.008. Mol Cell. 2017. PMID: 28306507 Free PMC article.
-
Epigenetic upregulation of metabotropic glutamate receptor 2 in the spinal cord attenuates oestrogen-induced visceral hypersensitivity.Gut. 2015 Dec;64(12):1913-20. doi: 10.1136/gutjnl-2014-307748. Epub 2014 Nov 6. Gut. 2015. PMID: 25378524 Free PMC article.
References
-
- Bodine PV, Harris HA, Komm BS. 1999. Suppression of ligand-dependent estrogen receptor activity by bone-resorbing cytokines in human osteoblasts. Endocrinology 140: 2439–2451 - PubMed
-
- Brower-Toland B, et al. 2005. Specific contributions of histone tails and their acetylation to the mechanical stability of nucleosomes. J. Mol. Biol. 346: 135–146 - PubMed
-
- Cakouros D, et al. 2001. A NF-kappa B/Sp1 region is essential for chromatin remodeling and correct transcription of a human granulocyte-macrophage colony-stimulating factor transgene. J. Immunol. 167: 302–310 - PubMed
-
- Carroll JS, et al. 2005. Chromosome-wide mapping of estrogen receptor binding reveals long-range regulation requiring the forkhead protein FoxA1. Cell 122: 33–43 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases