

Importance of the NCp7-like domain in the recognition of pre-let-7g by the pluripotency factor Lin28
- PMID: 22013165
- PMCID: PMC3287177
- DOI: 10.1093/nar/gkr808
Importance of the NCp7-like domain in the recognition of pre-let-7g by the pluripotency factor Lin28
Abstract
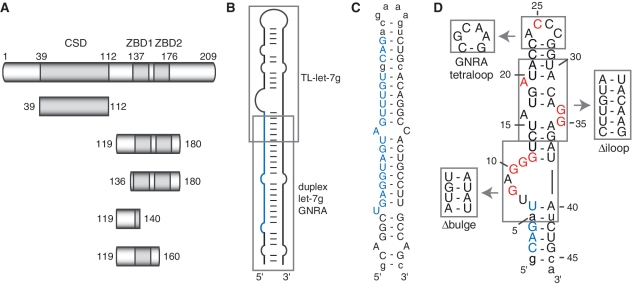
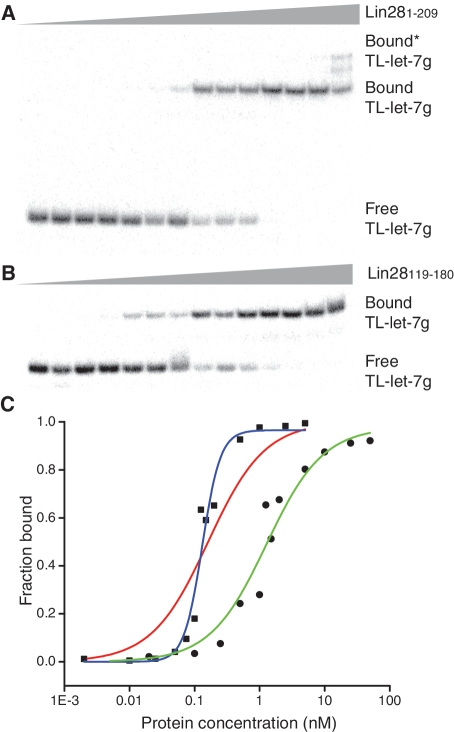
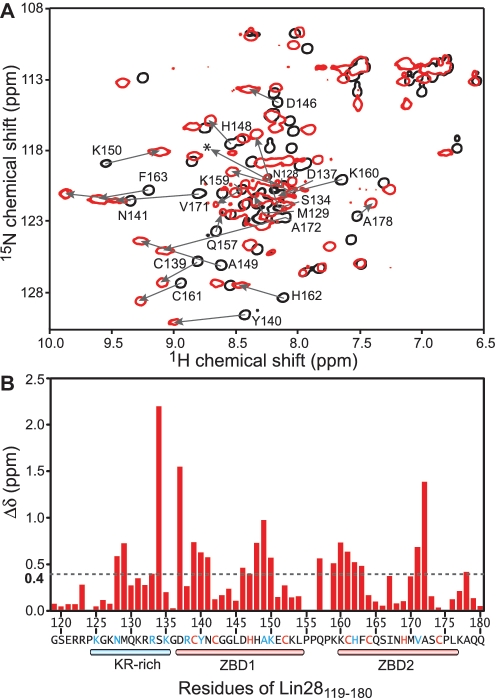
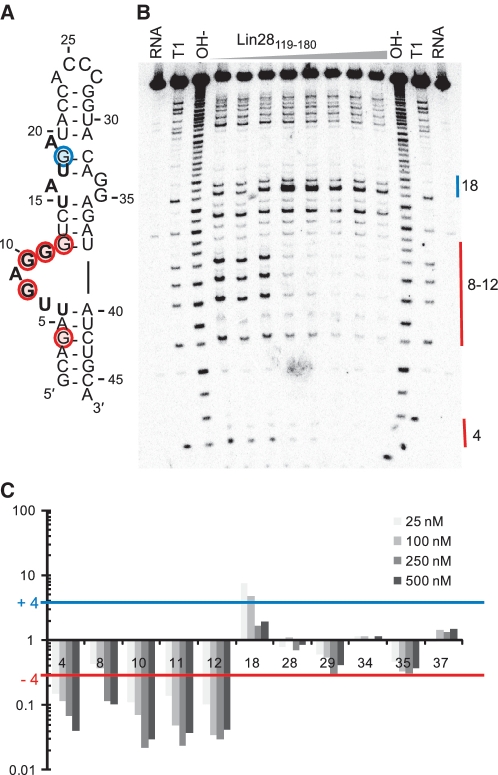
The pluripotency factor Lin28 is a highly conserved protein comprising a unique combination of RNA-binding motifs, an N-terminal cold-shock domain and a C-terminal region containing two retroviral-type CCHC zinc-binding domains. An important function of Lin28 is to inhibit the biogenesis of the let-7 family of microRNAs through a direct interaction with let-7 precursors. Here, we systematically characterize the determinants of the interaction between Lin28 and pre-let-7 g by investigating the effect of protein and RNA mutations on in vitro binding. We determine that Lin28 binds with high affinity to the extended loop of pre-let-7 g and that its C-terminal domain contributes predominantly to the affinity of this interaction. We uncover remarkable similarities between this C-terminal domain and the NCp7 protein of HIV-1, not only in terms of primary structure but also in their modes of RNA binding. This NCp7-like domain of Lin28 recognizes a G-rich bulge within pre-let-7 g, which is adjacent to one of the Dicer cleavage sites. We hypothesize that the NCp7-like domain initiates RNA binding and partially unfolds the RNA. This partial unfolding would then enable multiple copies of Lin28 to bind the extended loop of pre-let-7 g and protect the RNA from cleavage by the pre-microRNA processing enzyme Dicer.
Figures

Similar articles
-
Stepwise assembly of multiple Lin28 proteins on the terminal loop of let-7 miRNA precursors.Nucleic Acids Res. 2014 Apr;42(7):4615-28. doi: 10.1093/nar/gkt1391. Epub 2014 Jan 21. Nucleic Acids Res. 2014. PMID: 24452802 Free PMC article.
-
The Lin28 cold-shock domain remodels pre-let-7 microRNA.Nucleic Acids Res. 2012 Aug;40(15):7492-506. doi: 10.1093/nar/gks355. Epub 2012 May 8. Nucleic Acids Res. 2012. PMID: 22570413 Free PMC article.
-
A LIN28-dependent structural change in pre-let-7g directly inhibits dicer processing.Biochemistry. 2011 Sep 6;50(35):7514-21. doi: 10.1021/bi200851d. Epub 2011 Aug 11. Biochemistry. 2011. PMID: 21815640 Free PMC article.
-
Mechanisms of Lin28-mediated miRNA and mRNA regulation--a structural and functional perspective.Int J Mol Sci. 2013 Aug 9;14(8):16532-53. doi: 10.3390/ijms140816532. Int J Mol Sci. 2013. PMID: 23939427 Free PMC article. Review.
-
A mirror of two faces: Lin28 as a master regulator of both miRNA and mRNA.Wiley Interdiscip Rev RNA. 2012 Jul-Aug;3(4):483-94. doi: 10.1002/wrna.1112. Epub 2012 Mar 29. Wiley Interdiscip Rev RNA. 2012. PMID: 22467269 Review.
Cited by
-
Stepwise assembly of multiple Lin28 proteins on the terminal loop of let-7 miRNA precursors.Nucleic Acids Res. 2014 Apr;42(7):4615-28. doi: 10.1093/nar/gkt1391. Epub 2014 Jan 21. Nucleic Acids Res. 2014. PMID: 24452802 Free PMC article.
-
Short loop-targeting oligoribonucleotides antagonize Lin28 and enable pre-let-7 processing and suppression of cell growth in let-7-deficient cancer cells.Nucleic Acids Res. 2015 Jan;43(2):e9. doi: 10.1093/nar/gku1090. Epub 2014 Nov 6. Nucleic Acids Res. 2015. PMID: 25378324 Free PMC article.
-
Does Lin28 Antagonize miRNA-Mediated Repression by Displacing miRISC from Target mRNAs?Front Genet. 2012 Nov 16;3:240. doi: 10.3389/fgene.2012.00240. eCollection 2012. Front Genet. 2012. PMID: 23162570 Free PMC article.
-
MicroRNAs in HIV-1 infection: an integration of viral and cellular interaction at the genomic level.Front Microbiol. 2012 Aug 24;3:306. doi: 10.3389/fmicb.2012.00306. eCollection 2012. Front Microbiol. 2012. PMID: 22936931 Free PMC article.
-
Identification of LIN28B-bound mRNAs reveals features of target recognition and regulation.RNA Biol. 2013 Jul;10(7):1146-59. doi: 10.4161/rna.25194. Epub 2013 May 29. RNA Biol. 2013. PMID: 23770886 Free PMC article.
References
-
- Winter J, Jung S, Keller S, Gregory RI, Diederichs S. Many roads to maturity: microRNA biogenesis pathways and their regulation. Nat. Cell. Biol. 2009;11:228–234. - PubMed
-
- Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat. Rev. Genet. 2008;9:102–114. - PubMed
-
- Kim VN, Han J, Siomi MC. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009;10:126–139. - PubMed
-
- Zhao Y, Ransom JF, Li A, Vedantham V, von Drehle M, Muth AN, Tsuchihashi T, McManus MT, Schwartz RJ, Srivastava D. Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1-2. Cell. 2007;129:303–317. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
