

The RIG-I ATPase domain structure reveals insights into ATP-dependent antiviral signalling
- PMID: 21979817
- PMCID: PMC3207106
- DOI: 10.1038/embor.2011.190
The RIG-I ATPase domain structure reveals insights into ATP-dependent antiviral signalling
Abstract
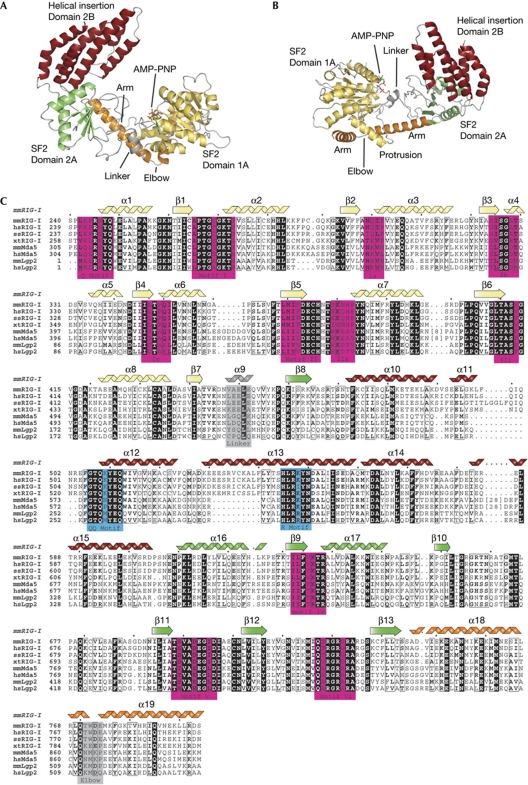
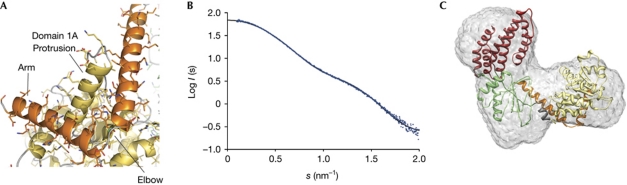
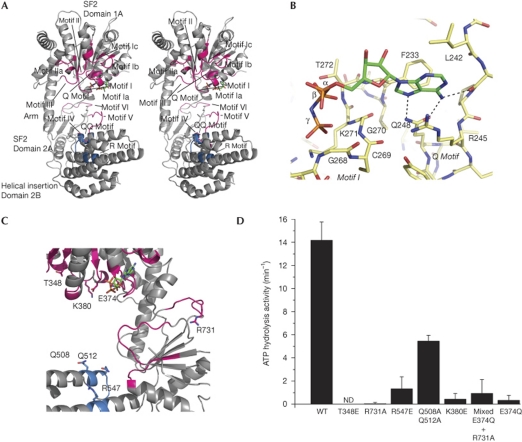
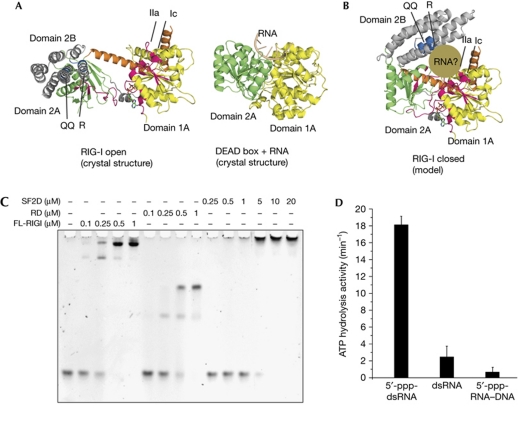
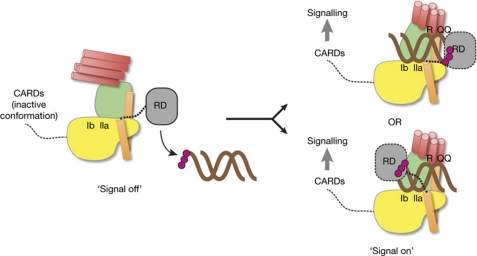
RIG-I detects cytosolic viral dsRNA with 5' triphosphates (5'-ppp-dsRNA), thereby initiating an antiviral innate immune response. Here we report the crystal structure of superfamily 2 (SF2) ATPase domain of RIG-I in complex with a nucleotide analogue. RIG-I SF2 comprises two RecA-like domains 1A and 2A and a helical insertion domain 2B, which together form a 'C'-shaped structure. Domains 1A and 2A are maintained in a 'signal-off' state with an inactive ATP hydrolysis site by an intriguing helical arm. By mutational analysis, we show surface motifs that are critical for dsRNA-stimulated ATPase activity, indicating that dsRNA induces a structural movement that brings domains 1A and 2A/B together to form an active ATPase site. The structure also indicates that the regulatory domain is close to the end of the helical arm, where it is well positioned to recruit 5'-ppp-dsRNA to the SF2 domain. Overall, our results indicate that the activation of RIG-I occurs through an RNA- and ATP-driven structural switch in the SF2 domain.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Structural insights into RNA recognition by RIG-I.Cell. 2011 Oct 14;147(2):409-22. doi: 10.1016/j.cell.2011.09.023. Cell. 2011. PMID: 22000018 Free PMC article.
-
Structural basis of RNA recognition and activation by innate immune receptor RIG-I.Nature. 2011 Sep 25;479(7373):423-7. doi: 10.1038/nature10537. Nature. 2011. PMID: 21947008 Free PMC article.
-
Kinetic discrimination of self/non-self RNA by the ATPase activity of RIG-I and MDA5.BMC Biol. 2015 Jul 28;13:54. doi: 10.1186/s12915-015-0166-9. BMC Biol. 2015. PMID: 26215161 Free PMC article.
-
Parts, assembly and operation of the RIG-I family of motors.Curr Opin Struct Biol. 2014 Apr;25:25-33. doi: 10.1016/j.sbi.2013.11.011. Epub 2013 Dec 20. Curr Opin Struct Biol. 2014. PMID: 24878341 Free PMC article. Review.
-
A structure-based model of RIG-I activation.RNA. 2012 Dec;18(12):2118-27. doi: 10.1261/rna.035949.112. Epub 2012 Nov 1. RNA. 2012. PMID: 23118418 Free PMC article. Review.
Cited by
-
An aromatic-rich loop couples DNA binding and ATP hydrolysis in the PriA DNA helicase.Nucleic Acids Res. 2016 Nov 16;44(20):9745-9757. doi: 10.1093/nar/gkw690. Epub 2016 Aug 2. Nucleic Acids Res. 2016. PMID: 27484483 Free PMC article.
-
Selective RNA targeting and regulated signaling by RIG-I is controlled by coordination of RNA and ATP binding.Nucleic Acids Res. 2017 Feb 17;45(3):1442-1454. doi: 10.1093/nar/gkw816. Nucleic Acids Res. 2017. PMID: 28180316 Free PMC article.
-
ATP hydrolysis by the viral RNA sensor RIG-I prevents unintentional recognition of self-RNA.Elife. 2015 Nov 26;4:e10859. doi: 10.7554/eLife.10859. Elife. 2015. PMID: 26609812 Free PMC article.
-
Molecular mechanics of RNA translocases.Methods Enzymol. 2012;511:131-47. doi: 10.1016/B978-0-12-396546-2.00006-1. Methods Enzymol. 2012. PMID: 22713318 Free PMC article.
-
Strategies of highly pathogenic RNA viruses to block dsRNA detection by RIG-I-like receptors: hide, mask, hit.Antiviral Res. 2013 Dec;100(3):615-35. doi: 10.1016/j.antiviral.2013.10.002. Epub 2013 Oct 12. Antiviral Res. 2013. PMID: 24129118 Free PMC article. Review.
References
-
- Cui S, Eisenacher K, Kirchhofer A, Brzozka K, Lammens A, Lammens K, Fujita T, Conzelmann KK, Krug A, Hopfner KP (2008) The C-terminal regulatory domain is the RNA 5′-triphosphate sensor of RIG-I. Mol Cell 29: 169–179 - PubMed
-
- Durr H, Korner C, Muller M, Hickmann V, Hopfner KP (2005) X-ray structures of the Sulfolobus solfataricus SWI2/SNF2 ATPase core and its complex with DNA. Cell 121: 363–373 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
