

The transformation of enterovirus replication structures: a three-dimensional study of single- and double-membrane compartments
- PMID: 21972238
- PMCID: PMC3187575
- DOI: 10.1128/mBio.00166-11
The transformation of enterovirus replication structures: a three-dimensional study of single- and double-membrane compartments
Abstract
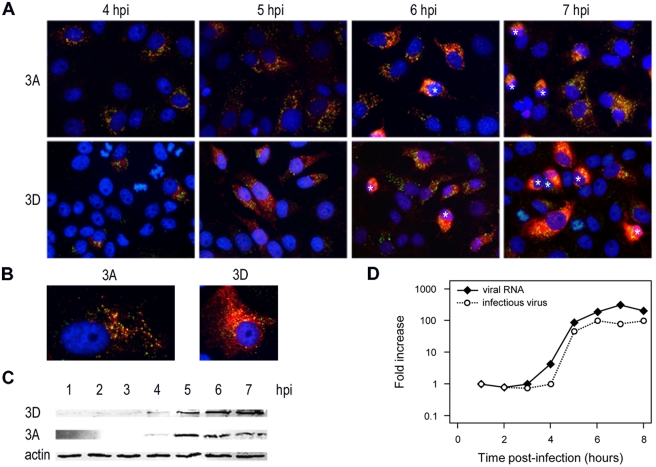
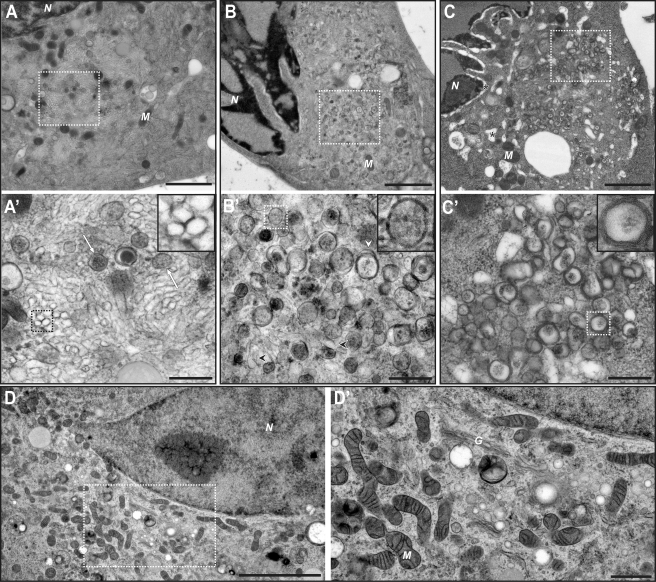
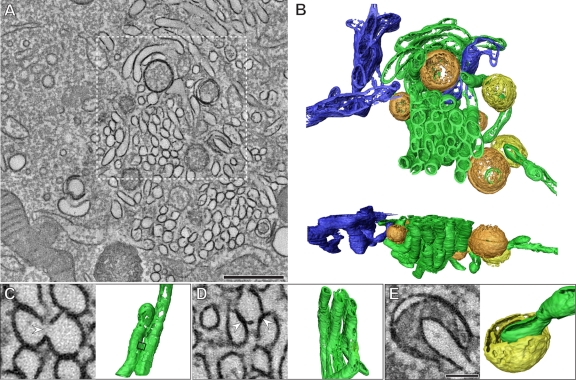
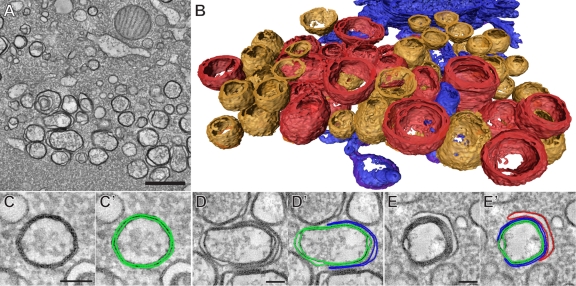
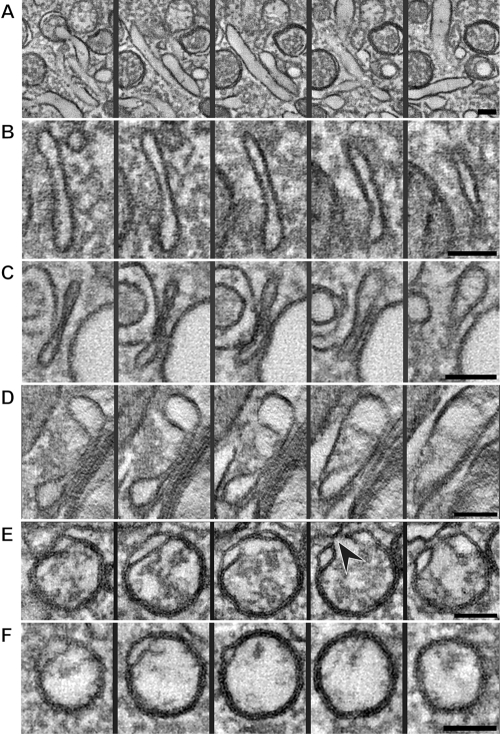
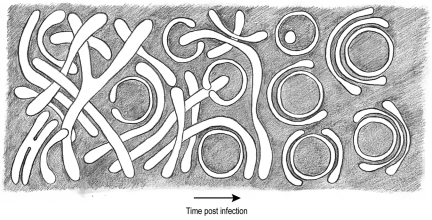
All positive-strand RNA viruses induce membrane structures in their host cells which are thought to serve as suitable microenvironments for viral RNA synthesis. The structures induced by enteroviruses, which are members of the family Picornaviridae, have so far been described as either single- or double-membrane vesicles (DMVs). Aside from the number of delimiting membranes, their exact architecture has also remained elusive due to the limitations of conventional electron microscopy. In this study, we used electron tomography (ET) to solve the three-dimensional (3-D) ultrastructure of these compartments. At different time points postinfection, coxsackievirus B3-infected cells were high-pressure frozen and freeze-substituted for ET analysis. The tomograms showed that during the exponential phase of viral RNA synthesis, closed smooth single-membrane tubules constituted the predominant virus-induced membrane structure, with a minor proportion of DMVs that were either closed or connected to the cytosol in a vase-like configuration. As infection progressed, the DMV number steadily increased, while the tubular single-membrane structures gradually disappeared. Late in infection, complex multilamellar structures, previously unreported, became apparent in the cytoplasm. Serial tomography disclosed that their basic unit is a DMV, which is enwrapped by one or multiple cisternae. ET also revealed striking intermediate structures that strongly support the conversion of single-membrane tubules into double-membrane and multilamellar structures by a process of membrane apposition, enwrapping, and fusion. Collectively, our work unravels the sequential appearance of distinct enterovirus-induced replication structures, elucidates their detailed 3-D architecture, and provides the basis for a model for their transformation during the course of infection.
Importance: Positive-strand RNA viruses hijack specific intracellular membranes and remodel them into special structures that support viral RNA synthesis. The ultrastructural characterization of these "replication structures" is key to understanding their precise role. Here, we resolved the three-dimensional architecture of enterovirus-induced membranous compartments and their transformation in time by applying electron tomography to cells infected with coxsackievirus B3 (CVB3). Our results show that closed single-membrane tubules are the predominant initial virus-induced structure, whereas double-membrane vesicles (DMVs) become increasingly abundant at the expense of these tubules as infection progresses. Additionally, more complex multilamellar structures appear late in infection. Based on compelling intermediate structures in our tomograms, we propose a model for transformation from the tubules to DMVs and multilamellar structures via enwrapping events. Our work provides an in-depth analysis of the development of an unsuspected variety of distinct replication structures during the course of CVB3 infection.
Figures
Similar articles
-
The Origin, Dynamic Morphology, and PI4P-Independent Formation of Encephalomyocarditis Virus Replication Organelles.mBio. 2018 Apr 17;9(2):e00420-18. doi: 10.1128/mBio.00420-18. mBio. 2018. PMID: 29666283 Free PMC article.
-
Ultrastructural Characterization of Turnip Mosaic Virus-Induced Cellular Rearrangements Reveals Membrane-Bound Viral Particles Accumulating in Vacuoles.J Virol. 2015 Dec;89(24):12441-56. doi: 10.1128/JVI.02138-15. Epub 2015 Sep 30. J Virol. 2015. PMID: 26423955 Free PMC article.
-
Infectious bronchitis virus generates spherules from zippered endoplasmic reticulum membranes.mBio. 2013 Oct 22;4(5):e00801-13. doi: 10.1128/mBio.00801-13. mBio. 2013. PMID: 24149513 Free PMC article.
-
Double-Membrane Vesicles as Platforms for Viral Replication.Trends Microbiol. 2020 Dec;28(12):1022-1033. doi: 10.1016/j.tim.2020.05.009. Epub 2020 Jun 11. Trends Microbiol. 2020. PMID: 32536523 Free PMC article. Review.
-
Biogenesis and architecture of arterivirus replication organelles.Virus Res. 2016 Jul 15;220:70-90. doi: 10.1016/j.virusres.2016.04.001. Epub 2016 Apr 9. Virus Res. 2016. PMID: 27071852 Free PMC article. Review.
Cited by
-
Antiviral Innate Immune Response Interferes with the Formation of Replication-Associated Membrane Structures Induced by a Positive-Strand RNA Virus.mBio. 2016 Dec 6;7(6):e01991-16. doi: 10.1128/mBio.01991-16. mBio. 2016. PMID: 27923923 Free PMC article.
-
Cryo-electron microscopy of nodavirus RNA replication organelles illuminates positive-strand RNA virus genome replication.Curr Opin Virol. 2021 Dec;51:74-79. doi: 10.1016/j.coviro.2021.09.008. Epub 2021 Sep 30. Curr Opin Virol. 2021. PMID: 34601307 Free PMC article. Review.
-
Expression and Cleavage of Middle East Respiratory Syndrome Coronavirus nsp3-4 Polyprotein Induce the Formation of Double-Membrane Vesicles That Mimic Those Associated with Coronaviral RNA Replication.mBio. 2017 Nov 21;8(6):e01658-17. doi: 10.1128/mBio.01658-17. mBio. 2017. PMID: 29162711 Free PMC article.
-
Membrane remodeling by SARS-CoV-2 - double-enveloped viral replication.Fac Rev. 2021 Feb 22;10:17. doi: 10.12703/r/10-17. eCollection 2021. Fac Rev. 2021. PMID: 33718934 Free PMC article. Review.
-
Enterovirus-D68 - A Reemerging Non-Polio Enterovirus that Causes Severe Respiratory and Neurological Disease in Children.Front Virol. 2024;4:1328457. doi: 10.3389/fviro.2024.1328457. Epub 2024 Feb 14. Front Virol. 2024. PMID: 39246649 Free PMC article.
References
-
- Caliguiri LA, Tamm I. 1969. Membranous structures associated with translation and transcription of poliovirus RNA. Science 166:885–886 - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
