

doi: 10.1038/nmeth.1714.
MS3 eliminates ratio distortion in isobaric multiplexed quantitative proteomics
Affiliations
- PMID: 21963607
- PMCID: PMC3205343
- DOI: 10.1038/nmeth.1714
Item in Clipboard
MS3 eliminates ratio distortion in isobaric multiplexed quantitative proteomics
Nat Methods.
.
Abstract
Quantitative mass spectrometry-based proteomics is highly versatile but not easily multiplexed. Isobaric labeling strategies allow mass spectrometry-based multiplexed proteome quantification; however, ratio distortion owing to protein quantification interference is a common effect. We present a two-proteome model (mixture of human and yeast proteins) in a sixplex isobaric labeling system to fully document the interference effect, and we report that applying triple-stage mass spectrometry (MS3) almost completely eliminates interference.
Figures
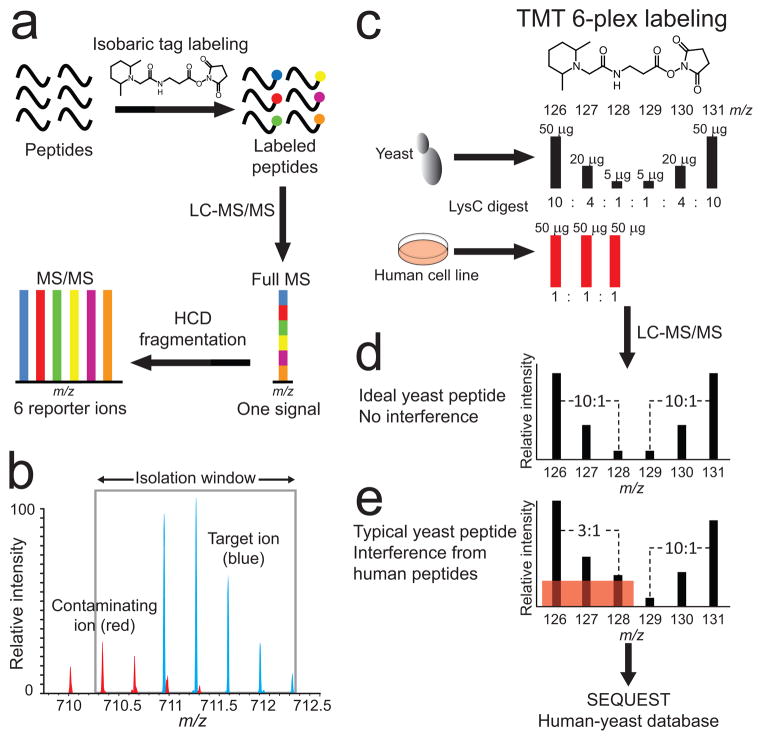
Isobaric labeling, interference and interference modeling. (a) Quantitative mass spectrometry-based proteomics experiments with stable isotope containing isobaric tags (TMT or iTRAQ) produce differentially labeled peptide ions that are indistinguishable in full-MS spectra. Quantification is based on low m/z reporter fragment ion intensities after a target ion is isolated and fragmented in an MS2 experiment. (b) Both accuracy and precision of peptide quantification are affected when contaminating peptide ions (red) are co-isolated with the target peptide ion (blue), a phenomenon that we term the interference effect. (c) Accurate measurement of the interference effect by a multi-proteome model using TMT six-plex labeled human cell line and S. cerevisiae Lys-C digests. The first three channels model interference, where yeast peptides were combined to create three different ratios (10:1, 4:1, 2.5:1) amongst unchanging interfering human peptides (1:1:1). In the last three channels, yeast peptides were combined in the same ratios (mirrored relative to the first three channels) without any human peptide interference. (d) An ideal yeast peptide MS2 spectrum without human peptide interference in the first three channels would have identical and mirrored TMT reporter ion intensities in the last three channels. (e) A typical yeast peptide has some degree of interference in the first three reporter ion channels (red shaded box), which results in ratio distortion towards 1:1:1.
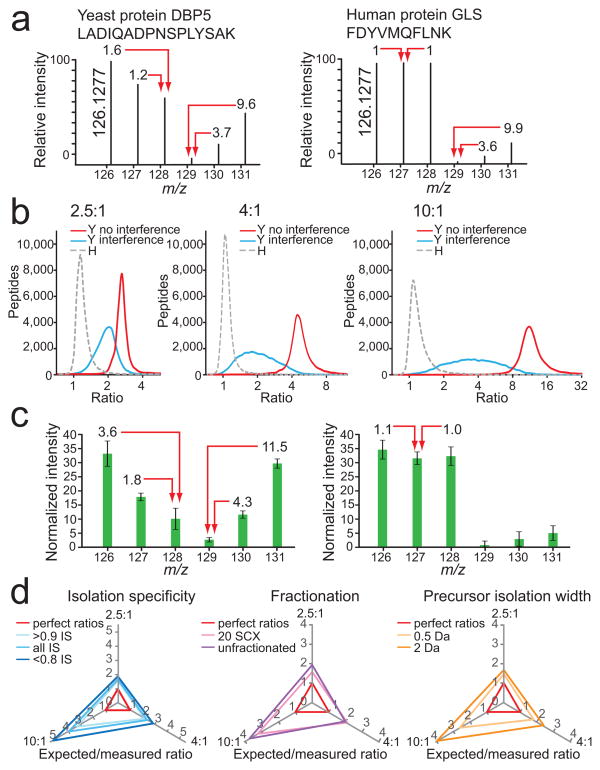
Evaluation and attempted removal of the interference effect. (a) An example of strong distortion of reporter ion intensities for a yeast peptide ion by interference of human peptide ions (left) and the effect of interference by yeast peptides on the reporter ion intensities of a human peptide (right; signals in channels 129, 130, and 131). (b) Ratiodistributions (log2) of yeast (Y) peptides in channels with human (H) peptide interference (blue line), without human peptide interference (red line), and human peptides only (gray dashed line) for the predicted ratios 2.5:1 (left), 4:1 (middle), and 10:1 (right). LC-MS2 data were collected after separating the yeast and human whole cell lysate digest into 20 fractions using SCX. (c) Averaged normalized relative intensities for each TMT reporter ion channel for yeast peptides (left), and human peptides (right) from the dataset described in (b). Error bars represent one standard deviation for 20,272 and 81,180 yeast and human peptides, respectively. (d) There was minimal improvement of yeast peptide ratios in channels with human peptide interference (126, 127, and 128) with post-acquisition data filtering of isolation specificity (IS; left), fractionation (middle) and precursor isolation width (right). Expected ratios were divided by measured ratios, where perfect ratios would have a value of 1 for all three axes (red triangle). The greater the distance of each tested condition from the red perfect ratio triangle, the greater the influence of human peptide interference on yeast peptide ratio distortion. Increasing dynamic range results in greater ratio distortion.
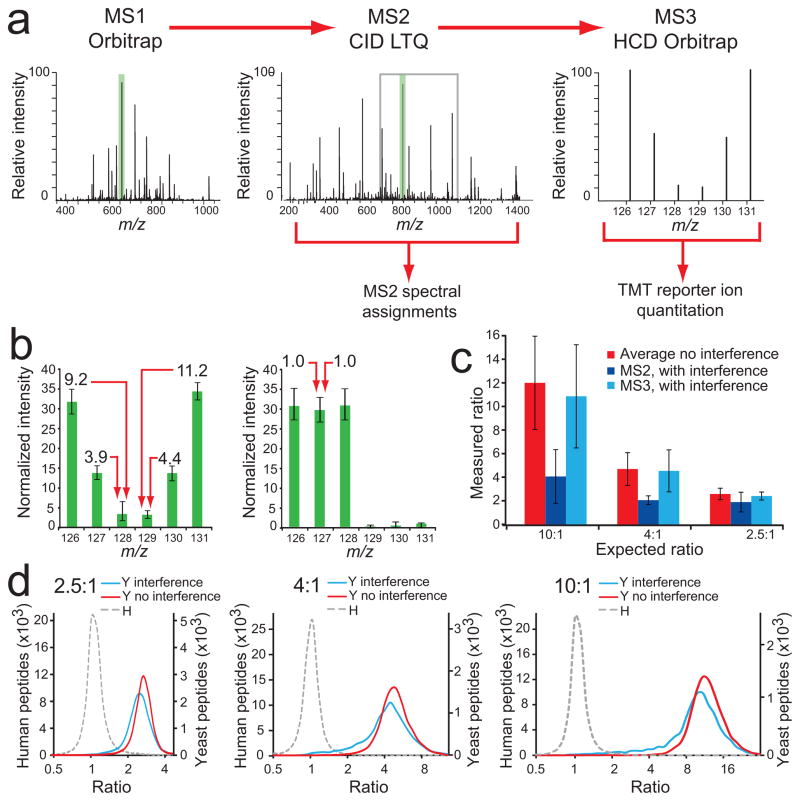
An MS3-based method eliminates the interference effect. (a) Our MS3 method separates quantification from peptide identification. Fast LTQ-CID-MS2 experiments are used for peptide sequence assignments, followed by the selection of the most intense fragment ion from MS2 (green shading) for HCD-MS3, within a 110–160% m/z range relative to the precursor ion m/z value (gray box), where TMT reporter ion intensities are measured in the Orbitrap. (b) Normalized intensities for each TMT reporter ion channel for yeast peptides (left), and human peptides (right). Almost no interference effect was measured, except for forward false positives (1.5%). Error bars represent one standard deviation for 8,919 and 65,595 yeast and human peptides, respectively (c) Yeast peptide ratios from dataset combining 20 SCX fractions. Ratios were substantially compressed in channels containing interference when quantification was based on MS2 spectra. Using MS3 for quantification, measured yeast peptide ratios from channels also containing human peptides (126, 127, 128) were very close to ratios measured for channels that contained only yeast peptide ions (129, 130, 131). Error bars represent one standard deviation for 29,813, 22,640 and 7,173 peptides for Average, MS2 and MS3 analyses, respectively. (d) Ratio distribution (log2 scale) of yeast (Y) peptides in channels with human (H) peptide interference (blue line), without human peptide interference (red line), and for human peptides (gray dashed line) for 2.5:1 (left), 4:1 (middle), and 10:1 (right) combinations.
Comment in
-
Taming the isobaric tagging elephant in the room in quantitative proteomics.Nat Methods. 2011 Oct 28;8(11):911-3. doi: 10.1038/nmeth.1736. Nat Methods. 2011. PMID: 22036744 No abstract available.
Similar articles
-
A Triple Knockout (TKO) Proteomics Standard for Diagnosing Ion Interference in Isobaric Labeling Experiments.J Am Soc Mass Spectrom. 2016 Oct;27(10):1620-5. doi: 10.1007/s13361-016-1434-9. Epub 2016 Jul 11. J Am Soc Mass Spectrom. 2016. PMID: 27400695 Free PMC article.
-
TKO6: A Peptide Standard To Assess Interference for Unit-Resolved Isobaric Labeling Platforms.J Proteome Res. 2019 Jan 4;18(1):565-570. doi: 10.1021/acs.jproteome.8b00902. Epub 2018 Dec 6. J Proteome Res. 2019. PMID: 30481031 Free PMC article.
-
High-resolution quadrupole improves spectral purity and reduces interference from non-target ions in isobaric multiplexed quantitative proteomics.Anal Chim Acta. 2024 Oct 9;1325:343135. doi: 10.1016/j.aca.2024.343135. Epub 2024 Aug 22. Anal Chim Acta. 2024. PMID: 39244297
-
Progress and pitfalls of using isobaric mass tags for proteome profiling.Expert Rev Proteomics. 2020 Feb;17(2):149-161. doi: 10.1080/14789450.2020.1731309. Epub 2020 Feb 20. Expert Rev Proteomics. 2020. PMID: 32067523 Review.
-
Development and application of proteomics technologies in Saccharomyces cerevisiae.Trends Biotechnol. 2005 Dec;23(12):598-604. doi: 10.1016/j.tibtech.2005.09.004. Epub 2005 Oct 3. Trends Biotechnol. 2005. PMID: 16202464 Review.
Cited by
-
Dual proteome-scale networks reveal cell-specific remodeling of the human interactome.Cell. 2021 May 27;184(11):3022-3040.e28. doi: 10.1016/j.cell.2021.04.011. Epub 2021 May 6. Cell. 2021. PMID: 33961781 Free PMC article.
-
Multisystem inflammatory syndrome in children is driven by zonulin-dependent loss of gut mucosal barrier.J Clin Invest. 2021 Jul 15;131(14):e149633. doi: 10.1172/JCI149633. J Clin Invest. 2021. PMID: 34032635 Free PMC article.
-
Quantifying proteomes and their post-translational modifications by stable isotope label-based mass spectrometry.Curr Opin Chem Biol. 2013 Oct;17(5):779-86. doi: 10.1016/j.cbpa.2013.06.011. Epub 2013 Jul 5. Curr Opin Chem Biol. 2013. PMID: 23835517 Free PMC article. Review.
-
Impact of eIF2α phosphorylation on the translational landscape of mouse embryonic stem cells.Cell Rep. 2024 Jan 23;43(1):113615. doi: 10.1016/j.celrep.2023.113615. Epub 2023 Dec 29. Cell Rep. 2024. PMID: 38159280 Free PMC article.
-
Mass Spectrometry-Based Approaches to Understand the Molecular Basis of Memory.Front Chem. 2016 Oct 14;4:40. doi: 10.3389/fchem.2016.00040. eCollection 2016. Front Chem. 2016. PMID: 27790611 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
