

Information transduction capacity of noisy biochemical signaling networks
- PMID: 21921160
- PMCID: PMC3895446
- DOI: 10.1126/science.1204553
Information transduction capacity of noisy biochemical signaling networks
Abstract
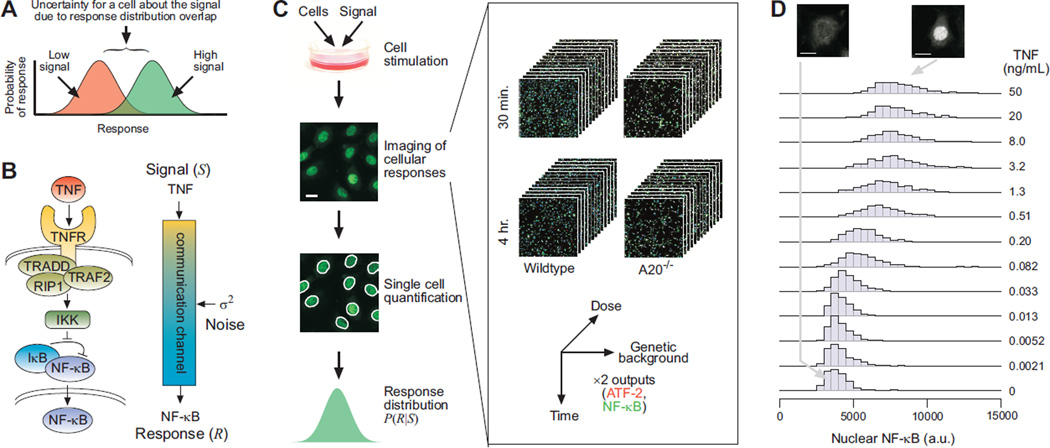
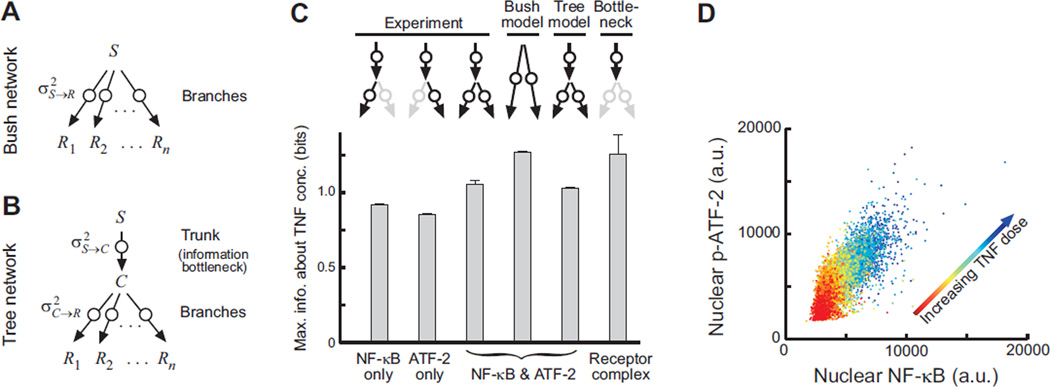
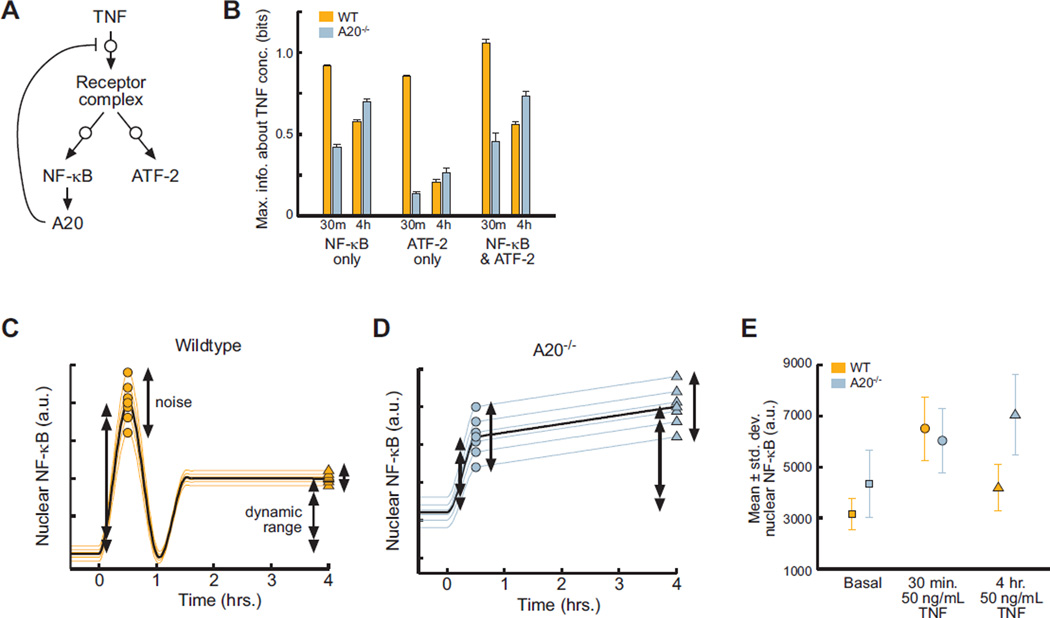
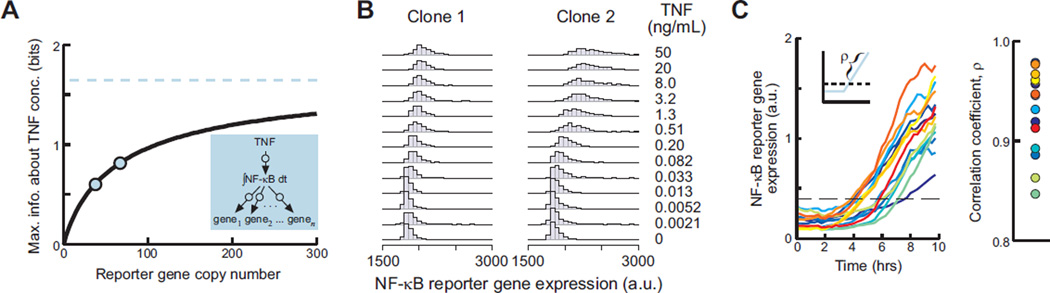
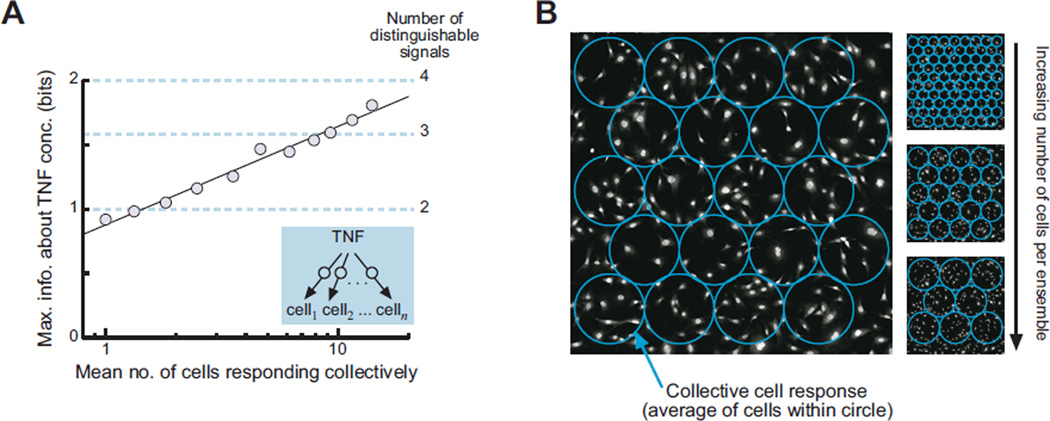
Molecular noise restricts the ability of an individual cell to resolve input signals of different strengths and gather information about the external environment. Transmitting information through complex signaling networks with redundancies can overcome this limitation. We developed an integrative theoretical and experimental framework, based on the formalism of information theory, to quantitatively predict and measure the amount of information transduced by molecular and cellular networks. Analyzing tumor necrosis factor (TNF) signaling revealed that individual TNF signaling pathways transduce information sufficient for accurate binary decisions, and an upstream bottleneck limits the information gained via multiple integrated pathways. Negative feedback to this bottleneck could both alleviate and enhance its limiting effect, despite decreasing noise. Bottlenecks likewise constrain information attained by networks signaling through multiple genes or cells.
Figures
Comment in
-
Cell signaling. Every bit counts.Science. 2011 Oct 21;334(6054):321-2. doi: 10.1126/science.1213834. Science. 2011. PMID: 22021849 No abstract available.
Similar articles
-
Noise decomposition of intracellular biochemical signaling networks using nonequivalent reporters.Proc Natl Acad Sci U S A. 2014 Dec 2;111(48):17330-5. doi: 10.1073/pnas.1411932111. Epub 2014 Nov 17. Proc Natl Acad Sci U S A. 2014. PMID: 25404303 Free PMC article.
-
Cell biology. Turning off inflammation signaling.Science. 2010 Feb 26;327(5969):1093-4. doi: 10.1126/science.1187271. Science. 2010. PMID: 20185718 No abstract available.
-
Encoding NF-kappaB temporal control in response to TNF: distinct roles for the negative regulators IkappaBalpha and A20.Genes Dev. 2008 Aug 1;22(15):2093-101. doi: 10.1101/gad.1680708. Genes Dev. 2008. PMID: 18676814 Free PMC article.
-
The ubiquitin-editing enzyme A20 (TNFAIP3) is a central regulator of immunopathology.Trends Immunol. 2009 Aug;30(8):383-91. doi: 10.1016/j.it.2009.05.007. Epub 2009 Jul 28. Trends Immunol. 2009. PMID: 19643665 Review.
-
Wires in the soup: quantitative models of cell signaling.Trends Cell Biol. 2008 Mar;18(3):112-8. doi: 10.1016/j.tcb.2008.01.005. Epub 2008 Feb 21. Trends Cell Biol. 2008. PMID: 18291655 Free PMC article. Review.
Cited by
-
Interplay of gene expression noise and ultrasensitive dynamics affects bacterial operon organization.PLoS Comput Biol. 2012;8(8):e1002672. doi: 10.1371/journal.pcbi.1002672. Epub 2012 Aug 30. PLoS Comput Biol. 2012. PMID: 22956903 Free PMC article.
-
Identifying sources of variation and the flow of information in biochemical networks.Proc Natl Acad Sci U S A. 2012 May 15;109(20):E1320-8. doi: 10.1073/pnas.1119407109. Epub 2012 Apr 23. Proc Natl Acad Sci U S A. 2012. PMID: 22529351 Free PMC article.
-
Distinct single-cell signaling characteristics are conferred by the MyD88 and TRIF pathways during TLR4 activation.Sci Signal. 2015 Jul 14;8(385):ra69. doi: 10.1126/scisignal.aaa5208. Sci Signal. 2015. PMID: 26175492 Free PMC article.
-
Structure and dynamics of molecular networks: a novel paradigm of drug discovery: a comprehensive review.Pharmacol Ther. 2013 Jun;138(3):333-408. doi: 10.1016/j.pharmthera.2013.01.016. Epub 2013 Feb 4. Pharmacol Ther. 2013. PMID: 23384594 Free PMC article. Review.
-
High cell density and high-resolution 3D bioprinting for fabricating vascularized tissues.Sci Adv. 2023 Feb 22;9(8):eade7923. doi: 10.1126/sciadv.ade7923. Epub 2023 Feb 22. Sci Adv. 2023. PMID: 36812321 Free PMC article.
References
-
- Rosenfeld N, Young JW, Alon U, Swain PS, Elowitz MB. Gene regulation at the single-cell level. Science. 2005;307:1962. - PubMed
-
- Blake WJ, Cantor KAMCR, Collins JJ. Noise in eukaryotic gene expression. Nature. 2003;422:633. - PubMed
-
- Elowitz MB, Levine AJ, Siggia ED, Swain PS. Stochastic gene expression in a single cell. Science. 2002;297:1183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
