

Insights into HLA-restricted T cell responses in a novel mouse model of dengue virus infection point toward new implications for vaccine design
- PMID: 21918184
- PMCID: PMC3186824
- DOI: 10.4049/jimmunol.1101970
Insights into HLA-restricted T cell responses in a novel mouse model of dengue virus infection point toward new implications for vaccine design
Abstract
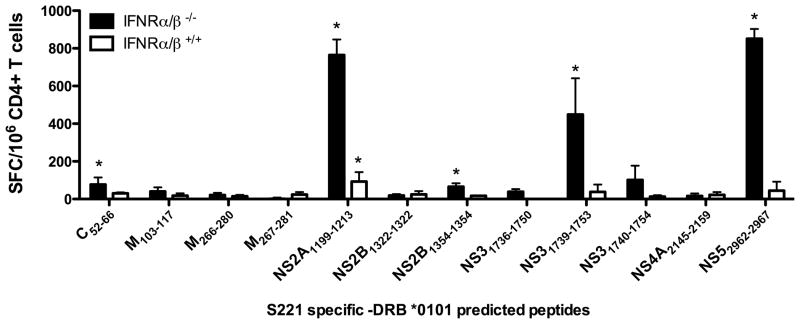
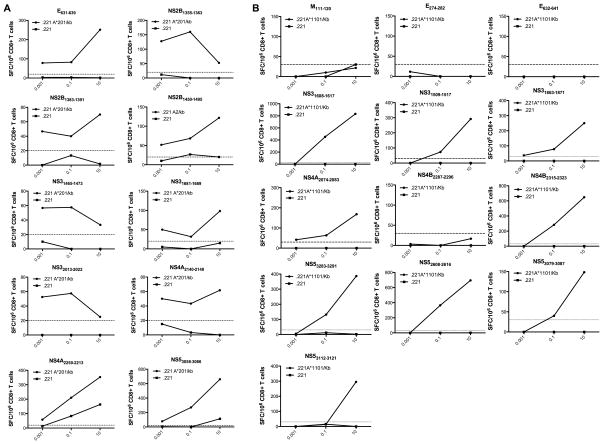
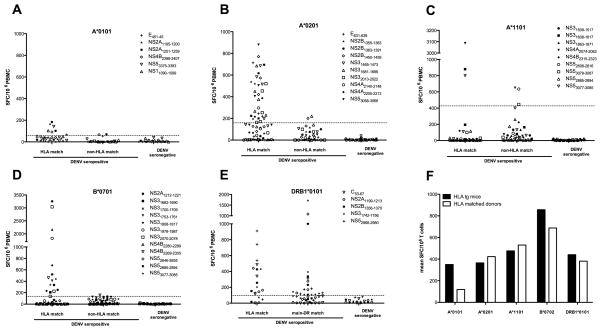
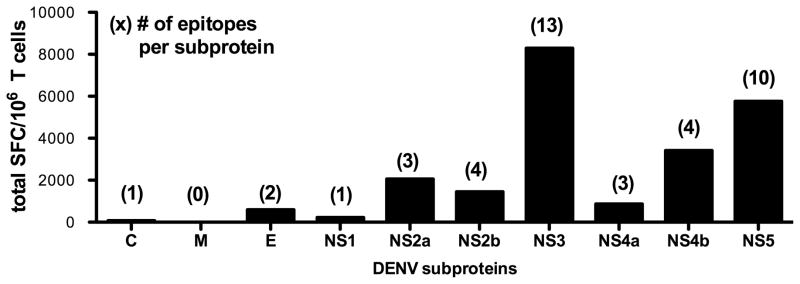
The frequency of dengue virus (DENV) infection has increased dramatically in the last few decades, and the lack of a vaccine has led to significant morbidity and mortality worldwide. To date, a convenient murine system to study human T cell responses to DENV has not been available. Mice transgenic for HLA are widely used to model human immune responses, and it has been shown that mouse-passaged DENV is able to replicate to significant levels in IFN-α/βR(-/-) mice. To cover a wide range of HLA phenotypes, we backcrossed IFN-α/βR(-/-) mice with HLA A*0201, A*0101, A*1101, B*0702, and DRB1*0101-transgenic mice. A DENV proteome-wide screen identified a total of 42 epitopes across all HLA-transgenic IFN-α/βR(-/-) strains tested. In contrast, only eight of these elicited responses in the corresponding IFN-α/βR(+/+) mice. We were able to identify T cell epitopes from 9 out of the 10 DENV proteins. However, the majority of responses were derived from the highly conserved nonstructural proteins NS3 and NS5. The relevance of this model is further demonstrated by the fact that most of the epitopes identified in our murine system are also recognized by PBMC from DENV-exposed human donors, and a dominance of HLA B*0702-restricted responses has been detected in both systems. Our results provide new insights into HLA-restricted T cell responses against DENV, and we describe in this study a novel murine model that allows the investigation of T cell-mediated immune mechanisms relevant to vaccine design.
Figures


Similar articles
-
A Modified mRNA Vaccine Targeting Immunodominant NS Epitopes Protects Against Dengue Virus Infection in HLA Class I Transgenic Mice.Front Immunol. 2019 Jun 21;10:1424. doi: 10.3389/fimmu.2019.01424. eCollection 2019. Front Immunol. 2019. PMID: 31293584 Free PMC article.
-
Identification of cytotoxic T lymphocyte epitopes in dengue virus serotype 1.J Med Virol. 2015 Jul;87(7):1077-89. doi: 10.1002/jmv.24167. Epub 2015 Mar 16. J Med Virol. 2015. PMID: 25777343
-
Identification of conserved and HLA-A*2402-restricted epitopes in Dengue virus serotype 2.Virus Res. 2015 Jan 22;196:5-12. doi: 10.1016/j.virusres.2014.10.022. Epub 2014 Nov 1. Virus Res. 2015. PMID: 25449574
-
Identification and selection of immunodominant B and T cell epitopes for dengue multi-epitope-based vaccine.Med Microbiol Immunol. 2021 Feb;210(1):1-11. doi: 10.1007/s00430-021-00700-x. Epub 2021 Jan 30. Med Microbiol Immunol. 2021. PMID: 33515283 Review.
-
Cross-Reactive T Cell Immunity to Dengue and Zika Viruses: New Insights Into Vaccine Development.Front Immunol. 2019 Jun 11;10:1316. doi: 10.3389/fimmu.2019.01316. eCollection 2019. Front Immunol. 2019. PMID: 31244855 Free PMC article. Review.
Cited by
-
Pathogenic Autoimmunity in Atherosclerosis Evolves From Initially Protective Apolipoprotein B100-Reactive CD4+ T-Regulatory Cells.Circulation. 2020 Sep 29;142(13):1279-1293. doi: 10.1161/CIRCULATIONAHA.119.042863. Epub 2020 Jul 24. Circulation. 2020. PMID: 32703007 Free PMC article.
-
Comprehensive analysis of dengue virus-specific responses supports an HLA-linked protective role for CD8+ T cells.Proc Natl Acad Sci U S A. 2013 May 28;110(22):E2046-53. doi: 10.1073/pnas.1305227110. Epub 2013 Apr 11. Proc Natl Acad Sci U S A. 2013. PMID: 23580623 Free PMC article.
-
Zika virus T-cell based 704/DNA vaccine promotes protection from Zika virus infection in the absence of neutralizing antibodies.PLoS Negl Trop Dis. 2024 Oct 17;18(10):e0012601. doi: 10.1371/journal.pntd.0012601. eCollection 2024 Oct. PLoS Negl Trop Dis. 2024. PMID: 39418312 Free PMC article.
-
T-cell immunity to infection with dengue virus in humans.Front Immunol. 2014 Mar 7;5:93. doi: 10.3389/fimmu.2014.00093. eCollection 2014. Front Immunol. 2014. PMID: 24639680 Free PMC article. Review.
-
CD8+ T cells mediate protection against Zika virus induced by an NS3-based vaccine.Sci Adv. 2020 Nov 4;6(45):eabb2154. doi: 10.1126/sciadv.abb2154. Print 2020 Nov. Sci Adv. 2020. PMID: 33148638 Free PMC article.
References
-
- Kyle JL, Harris E. Global spread and persistence of dengue. Annu Rev Microbiol. 2008;62:71–92. - PubMed
-
- WHO. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control. Geneva, Switzerland: 2009. - PubMed
-
- Burke D, Monath T. Flaviviruses. In: Knipe aPHD., editor. Field’s Virology. William and Wilkins; Philadelphia: 2001. pp. 1043–1126.
-
- Rothman AL. Cellular immunology of sequential dengue virus infection and its role in disease pathogenesis. Curr Top Microbiol Immunol. 2010;338:83–98. - PubMed
-
- Mathew A, Rothman AL. Understanding the contribution of cellular immunity to dengue disease pathogenesis. Immunol Rev. 2008;225:300–313. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials