

Regulation of the Fanconi anemia pathway by a SUMO-like delivery network
- PMID: 21896657
- PMCID: PMC3175720
- DOI: 10.1101/gad.17020911
Regulation of the Fanconi anemia pathway by a SUMO-like delivery network
Abstract
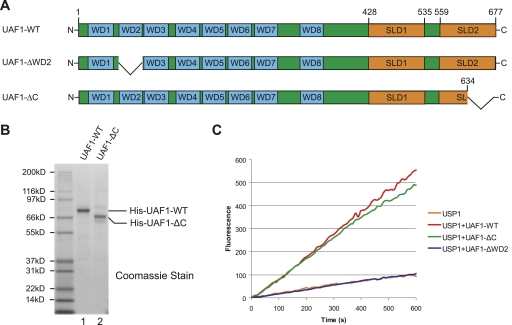
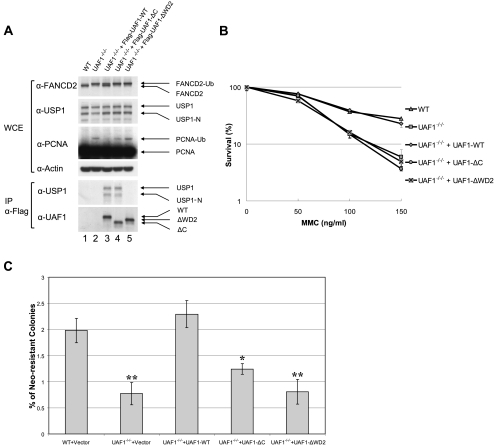
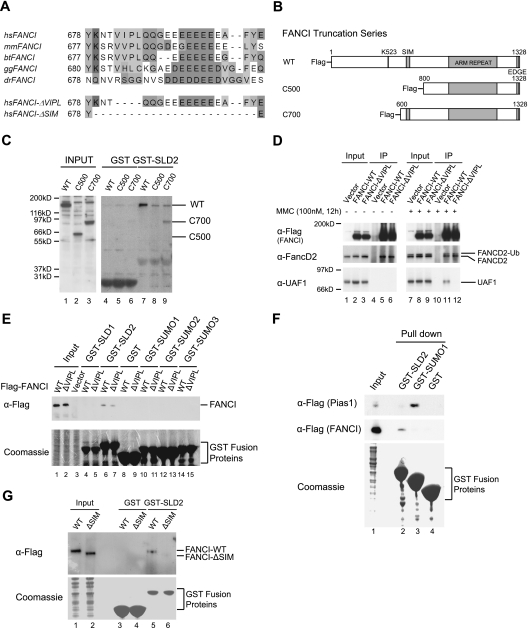
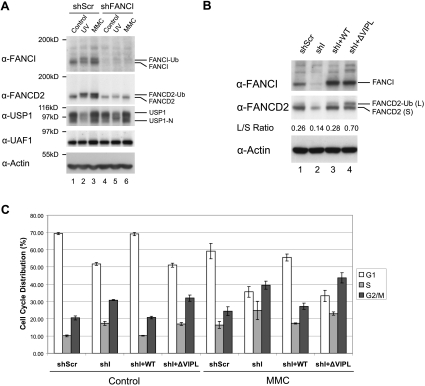
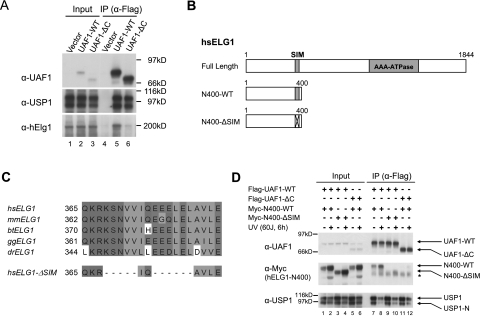
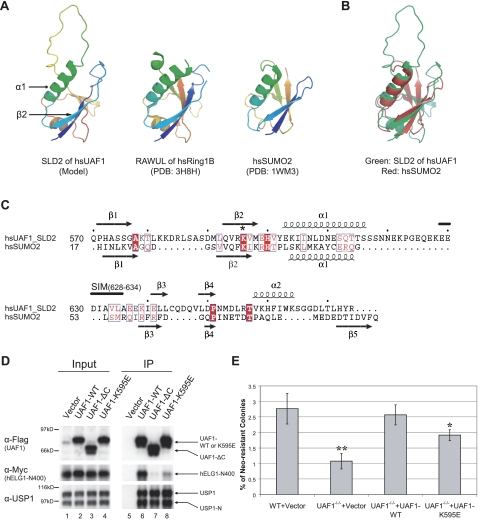
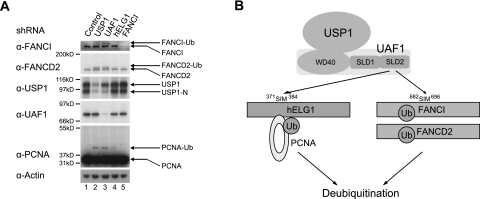
The USP1/UAF1 complex deubiquitinates the Fanconi anemia protein FANCD2, thereby promoting homologous recombination and DNA cross-link repair. How USP1/UAF1 is targeted to the FANCD2/FANCI heterodimer has remained unknown. Here we show that UAF1 contains a tandem repeat of SUMO-like domains in its C terminus (SLD1 and SLD2). SLD2 binds directly to a SUMO-like domain-interacting motif (SIM) on FANCI. Deletion of the SLD2 sequence of UAF1 or mutation of the SIM on FANCI disrupts UAF1/FANCI binding and inhibits FANCD2 deubiquitination and DNA repair. The USP1/UAF1 complex also deubiquitinates PCNA-Ub, and deubiquitination requires the PCNA-binding protein hELG1. The SLD2 sequence of UAF1 binds to a SIM on hELG1, thus targeting the USP1/UAF1 complex to its PCNA-Ub substrate. We propose that the regulated targeting of USP1/UAF1 to its DNA repair substrates, FANCD2-Ub and PCNA-Ub, by SLD-SIM interactions coordinates homologous recombination and translesion DNA synthesis.
Figures
Comment in
-
Shared and unique properties of ubiquitin and SUMO interaction networks in DNA repair.Genes Dev. 2011 Sep 1;25(17):1763-9. doi: 10.1101/gad.17593511. Genes Dev. 2011. PMID: 21896653 Free PMC article.
Similar articles
-
The Fanconi anemia ID2 complex: dueling saxes at the crossroads.Cell Cycle. 2014;13(19):2999-3015. doi: 10.4161/15384101.2014.956475. Cell Cycle. 2014. PMID: 25486561 Free PMC article. Review.
-
Human ELG1 regulates the level of ubiquitinated proliferating cell nuclear antigen (PCNA) through Its interactions with PCNA and USP1.J Biol Chem. 2010 Apr 2;285(14):10362-9. doi: 10.1074/jbc.M109.092544. Epub 2010 Feb 9. J Biol Chem. 2010. PMID: 20147293 Free PMC article.
-
The USP1/UAF1 complex promotes double-strand break repair through homologous recombination.Mol Cell Biol. 2011 Jun;31(12):2462-9. doi: 10.1128/MCB.05058-11. Epub 2011 Apr 11. Mol Cell Biol. 2011. PMID: 21482670 Free PMC article.
-
Structural basis of FANCD2 deubiquitination by USP1-UAF1.Nat Struct Mol Biol. 2021 Apr;28(4):356-364. doi: 10.1038/s41594-021-00576-8. Epub 2021 Apr 1. Nat Struct Mol Biol. 2021. PMID: 33795880
-
Mechanism, specificity, and function of FANCD2-FANCI ubiquitination and deubiquitination.FEBS J. 2022 Aug;289(16):4811-4829. doi: 10.1111/febs.16077. Epub 2021 Jun 29. FEBS J. 2022. PMID: 34137174 Review.
Cited by
-
ATAD5 restricts R-loop formation through PCNA unloading and RNA helicase maintenance at the replication fork.Nucleic Acids Res. 2020 Jul 27;48(13):7218-7238. doi: 10.1093/nar/gkaa501. Nucleic Acids Res. 2020. PMID: 32542338 Free PMC article.
-
Efficient embryonic homozygous gene conversion via RAD51-enhanced interhomolog repair.Cell. 2021 Jun 10;184(12):3267-3280.e18. doi: 10.1016/j.cell.2021.04.035. Epub 2021 May 26. Cell. 2021. PMID: 34043941 Free PMC article.
-
SUMO rules: regulatory concepts and their implication in neurologic functions.Neuromolecular Med. 2013 Dec;15(4):639-60. doi: 10.1007/s12017-013-8258-6. Epub 2013 Aug 30. Neuromolecular Med. 2013. PMID: 23990202 Review.
-
Rescue of replication failure by Fanconi anaemia proteins.Chromosoma. 2012 Feb;121(1):21-36. doi: 10.1007/s00412-011-0349-2. Epub 2011 Nov 6. Chromosoma. 2012. PMID: 22057367 Free PMC article. Review.
-
Insert L1 is a central hub for allosteric regulation of USP1 activity.EMBO Rep. 2021 Apr 7;22(4):e51749. doi: 10.15252/embr.202051749. Epub 2021 Feb 23. EMBO Rep. 2021. PMID: 33619839 Free PMC article.
References
-
- Bergink S, Jentsch S 2009. Principles of ubiquitin and SUMO modifications in DNA repair. Nature 458: 461–467 - PubMed
-
- Bezsonova I, Walker JR, Bacik JP, Duan S, Dhe-Paganon S, Arrowsmith CH 2009. Ring1B contains a ubiquitin-like docking module for interaction with Cbx proteins. Biochemistry 48: 10542–10548 - PubMed
-
- Bienko M, Green CM, Sabbioneda S, Crosetto N, Matic I, Hibbert RG, Begovic T, Niimi A, Mann M, Lehmann AR, et al. 2010. Regulation of translesion synthesis DNA polymerase eta by monoubiquitination. Mol Cell 37: 396–407 - PubMed
-
- Cohn MA, Kowal P, Yang K, Haas W, Huang TT, Gygi SP, D'Andrea AD 2007. A UAF1-containing multisubunit protein complex regulates the Fanconi anemia pathway. Mol Cell 28: 786–797 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous