

Deletion of cannabinoid receptors 1 and 2 exacerbates APC function to increase inflammation and cellular immunity during influenza infection
- PMID: 21873455
- PMCID: PMC3206470
- DOI: 10.1189/jlb.0511219
Deletion of cannabinoid receptors 1 and 2 exacerbates APC function to increase inflammation and cellular immunity during influenza infection
Abstract
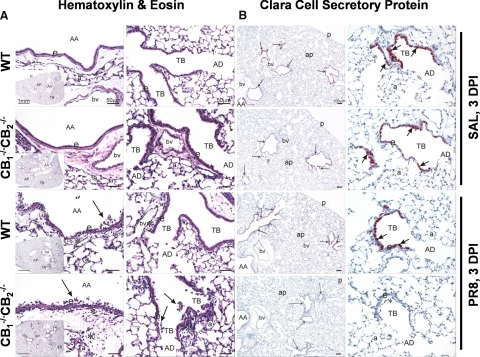
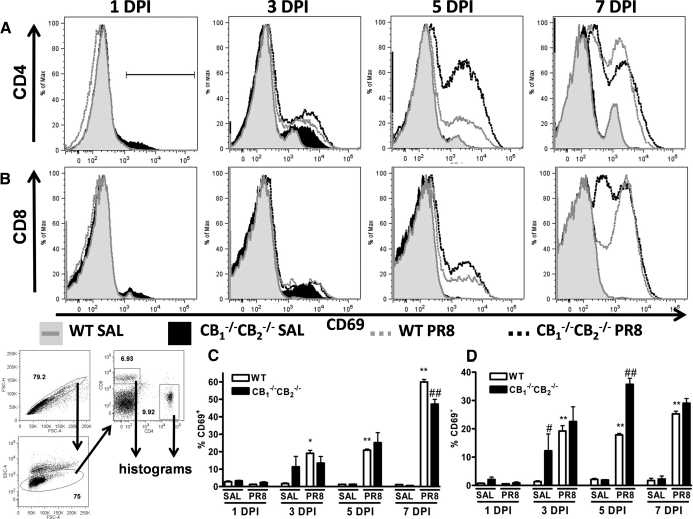
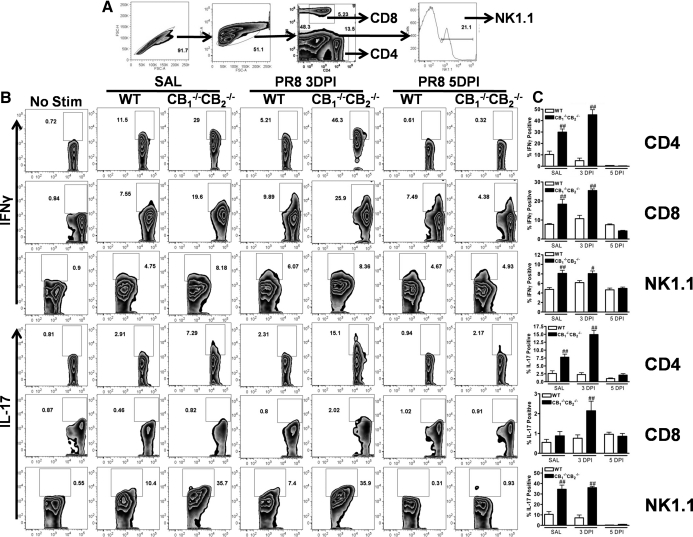
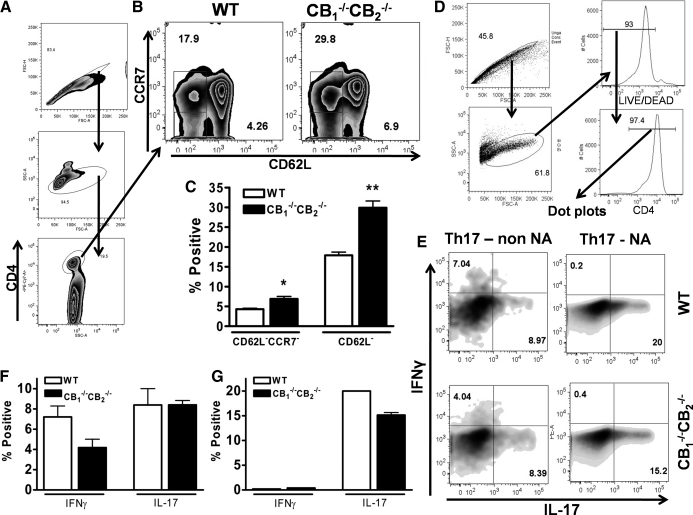
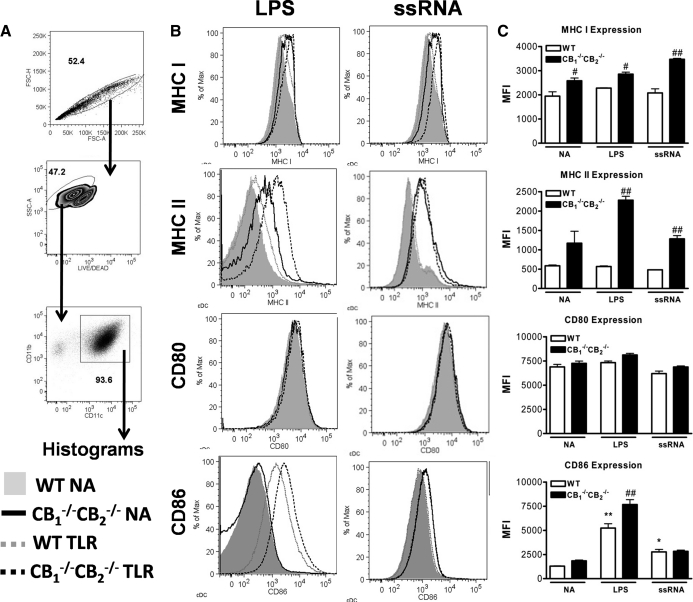
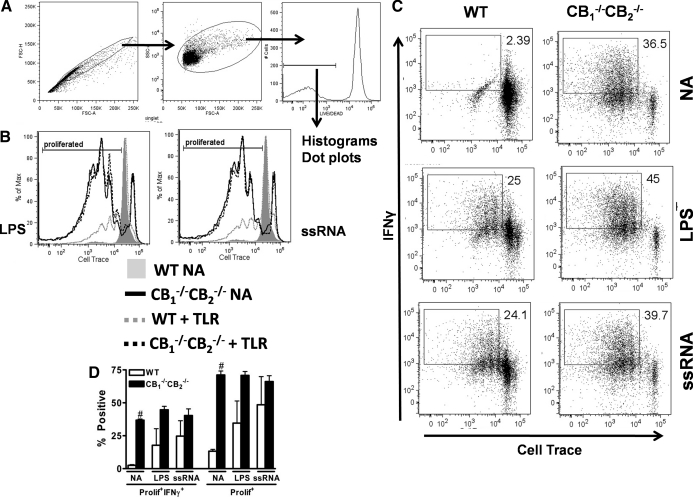
We and others have reported that simultaneous targeted deletion of CB(1) and CB(2) resulted in exacerbation of immune reactivity, suggesting a role of endocannabinoids in down-regulating immune function. In this study, we demonstrate that APC function is enhanced specifically in the absence of CB(1) and CB(2) signaling, resulting in an exacerbated immune response phenotype. After influenza infection, CB(1)(-/-)CB(2)(-/-) mice showed more pronounced pulmonary damage, increased inflammatory cell infiltrate, inflammation, and a greater cellular immune responses compared with WT mice, as evidenced by transcriptome analysis, more robust T cell activation, and effector cell cytokine production. After direct activation in vitro, there were no differences in the percentages of cytokine-producing CD4(+) T cells between CB(1)(-/-)CB(2)(-/-) and WT mice. However, untreated CB(1)(-/-)CB(2)(-/-) mice routinely had fewer naïve T cells compared with WT, suggesting dysregulation of APC immune homeostasis. Moreover, bmDCs and AM isolated from CB(1)(-/-)CB(2)(-/-) mice exhibited a more mature phenotype, with and without TLR stimulation, and bmDCs elicited T cells more robustly than WT mice. Collectively, these findings implicate a role for CB(1) and CB(2) on APCs in regulating immune responses and immune homeostasis.
Figures

Similar articles
-
Targeted deletion of cannabinoid receptors CB1 and CB2 produced enhanced inflammatory responses to influenza A/PR/8/34 in the absence and presence of Delta9-tetrahydrocannabinol.J Leukoc Biol. 2008 Mar;83(3):785-96. doi: 10.1189/jlb.0907618. Epub 2007 Dec 11. J Leukoc Biol. 2008. PMID: 18073275
-
Δ9-tetrahydrocannabinol impairs the inflammatory response to influenza infection: role of antigen-presenting cells and the cannabinoid receptors 1 and 2.Toxicol Sci. 2013 Feb;131(2):419-33. doi: 10.1093/toxsci/kfs315. Epub 2012 Nov 14. Toxicol Sci. 2013. PMID: 23152191 Free PMC article.
-
The effects of targeted deletion of cannabinoid receptors CB1 and CB2 on intranasal sensitization and challenge with adjuvant-free ovalbumin.Toxicol Pathol. 2010 Apr;38(3):382-92. doi: 10.1177/0192623310362706. Epub 2010 Feb 26. Toxicol Pathol. 2010. PMID: 20190202 Free PMC article.
-
The role of CB1 in immune modulation by cannabinoids.Pharmacol Ther. 2013 Mar;137(3):365-74. doi: 10.1016/j.pharmthera.2012.12.004. Epub 2012 Dec 20. Pharmacol Ther. 2013. PMID: 23261520 Review.
-
Endocannabinoids and the Immune System in Health and Disease.Handb Exp Pharmacol. 2015;231:185-211. doi: 10.1007/978-3-319-20825-1_6. Handb Exp Pharmacol. 2015. PMID: 26408161 Review.
Cited by
-
Effects of Cannabinoids on T-cell Function and Resistance to Infection.J Neuroimmune Pharmacol. 2015 Jun;10(2):204-16. doi: 10.1007/s11481-015-9603-3. Epub 2015 Apr 16. J Neuroimmune Pharmacol. 2015. PMID: 25876735 Free PMC article. Review.
-
Δ9-Tetrahydrocannabinol Suppresses Monocyte-Mediated Astrocyte Production of Monocyte Chemoattractant Protein 1 and Interleukin-6 in a Toll-Like Receptor 7-Stimulated Human Coculture.J Pharmacol Exp Ther. 2019 Oct;371(1):191-201. doi: 10.1124/jpet.119.260661. Epub 2019 Aug 5. J Pharmacol Exp Ther. 2019. PMID: 31383729 Free PMC article.
-
Enhanced humoral immunity in mice lacking CB1 and CB2 receptors (Cnr1-/-/Cnr2-/- mice) is not due to increased splenic noradrenergic neuronal activity.J Neuroimmune Pharmacol. 2014 Sep;9(4):544-57. doi: 10.1007/s11481-014-9549-x. Epub 2014 May 29. J Neuroimmune Pharmacol. 2014. PMID: 24870806
-
Exposure to Δ9-Tetrahydrocannabinol Impairs the Differentiation of Human Monocyte-derived Dendritic Cells and their Capacity for T cell Activation.J Neuroimmune Pharmacol. 2015 Jun;10(2):333-43. doi: 10.1007/s11481-015-9587-z. Epub 2015 Jan 23. J Neuroimmune Pharmacol. 2015. PMID: 25614186 Free PMC article.
-
Characterization of endocannabinoid-mediated induction of myeloid-derived suppressor cells involving mast cells and MCP-1.J Leukoc Biol. 2014 Apr;95(4):609-19. doi: 10.1189/jlb.0613350. Epub 2013 Dec 6. J Leukoc Biol. 2014. PMID: 24319288 Free PMC article.
References
-
- Holt P. G., Strickland D. H., Wikstrom M. E., Jahnsen F. L. (2008) Regulation of immunological homeostasis in the respiratory tract. Nat. Rev. Immunol. 8, 142–152 - PubMed
-
- Kobayashi K. S., Flavell R. A. (2004) Shielding the double-edged sword: negative regulation of the innate immune system. J. Leukoc. Biol. 75, 428–433 - PubMed
-
- Wong P., Pamer E. G. (2003) Feedback regulation of pathogen-specific T cell priming. Immunity 18, 499–511 - PubMed
-
- Banchereau J., Steinman R. M. (1998) Dendritic cells and the control of immunity. Nature 392, 245–252 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
