

Small-molecule activators of TMEM16A, a calcium-activated chloride channel, stimulate epithelial chloride secretion and intestinal contraction
- PMID: 21836025
- PMCID: PMC3205834
- DOI: 10.1096/fj.11-191627
Small-molecule activators of TMEM16A, a calcium-activated chloride channel, stimulate epithelial chloride secretion and intestinal contraction
Abstract
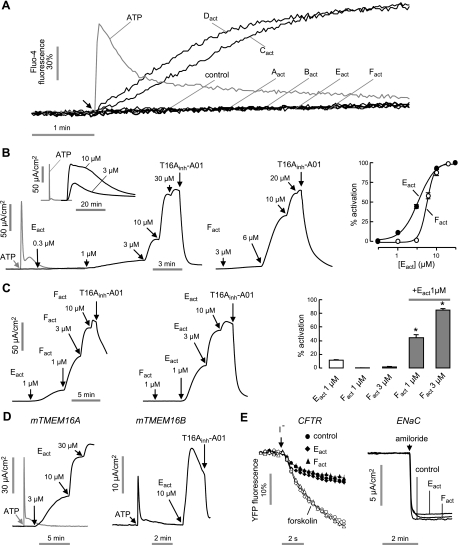
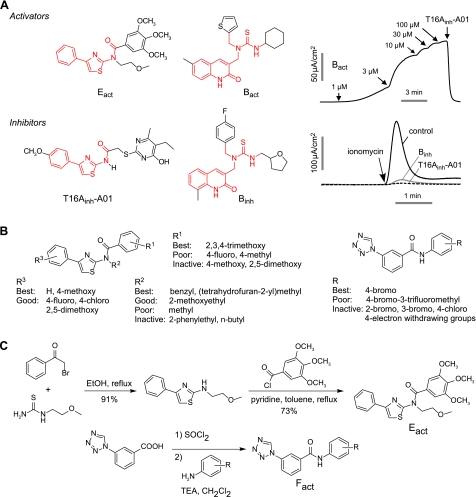
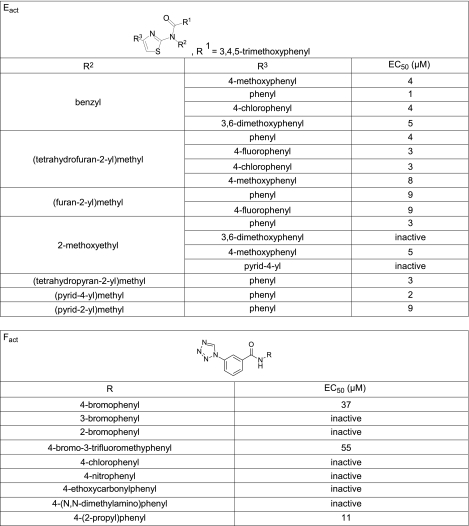
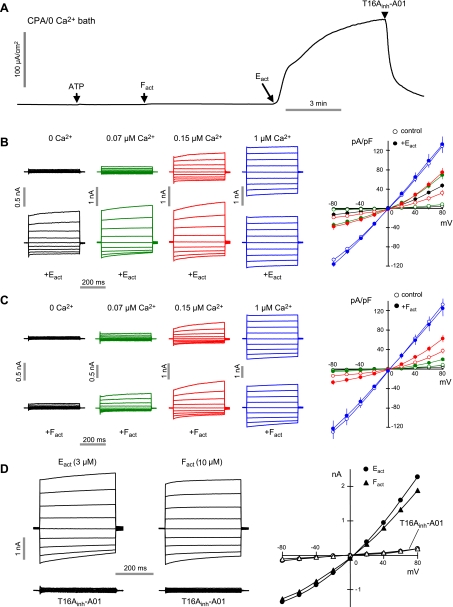
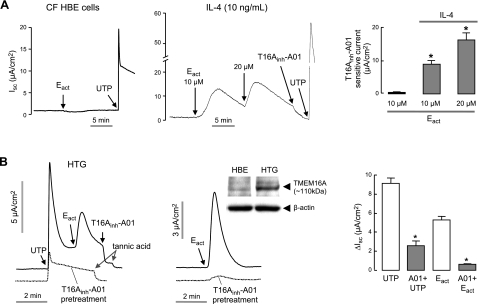
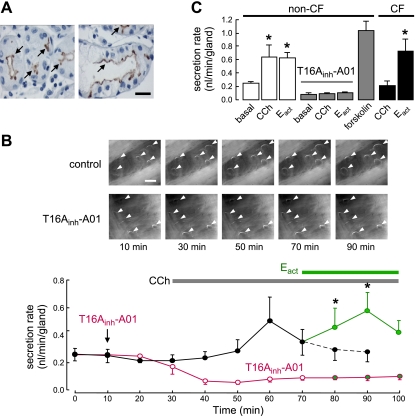
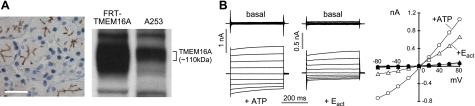
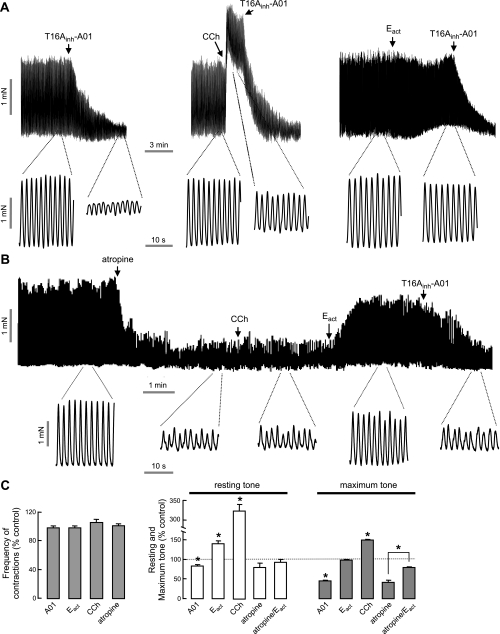
TMEM16A (ANO1) is a calcium-activated chloride channel (CaCC) expressed in secretory epithelia, smooth muscle, and other tissues. Cell-based functional screening of ∼110,000 compounds revealed compounds that activated TMEM16A CaCC conductance without increasing cytoplasmic Ca(2+). By patch-clamp, N-aroylaminothiazole "activators" (E(act)) strongly increased Cl(-) current at 0 Ca(2+), whereas tetrazolylbenzamide "potentiators" (F(act)) were not active at 0 Ca(2+) but reduced the EC(50) for Ca(2+)-dependent TMEM16A activation. Of 682 analogs tested, the most potent activator (E(act)) and potentiator (F(act)) produced large and more sustained CaCC Cl(-) currents than general agonists of Ca(2+) signaling, with EC(50) 3-6 μM and Cl(-) conductance comparable to that induced transiently by Ca(2+)-elevating purinergic agonists. Analogs of activators were identified that fully inhibited TMEM16A Cl(-) conductance, providing further evidence for direct TMEM16A binding. The TMEM16A activators increased CaCC conductance in human salivary and airway submucosal gland epithelial cells, and IL-4 treated bronchial cells, and stimulated submucosal gland secretion in human bronchi and smooth muscle contraction in mouse intestine. Small-molecule, TMEM16A-targeted activators may be useful for drug therapy of cystic fibrosis, dry mouth, and gastrointestinal hypomotility disorders, and for pharmacological dissection of TMEM16A function.
Figures

Similar articles
-
TMEM16A inhibitors reveal TMEM16A as a minor component of calcium-activated chloride channel conductance in airway and intestinal epithelial cells.J Biol Chem. 2011 Jan 21;286(3):2365-74. doi: 10.1074/jbc.M110.175109. Epub 2010 Nov 17. J Biol Chem. 2011. PMID: 21084298 Free PMC article.
-
Control of TMEM16A by INO-4995 and other inositolphosphates.Br J Pharmacol. 2013 Jan;168(1):253-65. doi: 10.1111/j.1476-5381.2012.02193.x. Br J Pharmacol. 2013. PMID: 22946960 Free PMC article.
-
Revealing the activation pathway for TMEM16A chloride channels from macroscopic currents and kinetic models.Pflugers Arch. 2016 Jul;468(7):1241-1257. doi: 10.1007/s00424-016-1830-9. Epub 2016 May 2. Pflugers Arch. 2016. PMID: 27138167 Free PMC article.
-
Calcium-Activated Cl- Channel: Insights on the Molecular Identity in Epithelial Tissues.Int J Mol Sci. 2018 May 10;19(5):1432. doi: 10.3390/ijms19051432. Int J Mol Sci. 2018. PMID: 29748496 Free PMC article. Review.
-
Bestrophin and TMEM16-Ca(2+) activated Cl(-) channels with different functions.Cell Calcium. 2009 Oct;46(4):233-41. doi: 10.1016/j.ceca.2009.09.003. Epub 2009 Sep 26. Cell Calcium. 2009. PMID: 19783045 Review.
Cited by
-
The molecular basis of the genesis of basal tone in internal anal sphincter.Nat Commun. 2016 Apr 22;7:11358. doi: 10.1038/ncomms11358. Nat Commun. 2016. PMID: 27101932 Free PMC article.
-
Anion efflux mediates transduction in the hair cells of the zebrafish lateral line.Proc Natl Acad Sci U S A. 2023 Dec 26;120(52):e2315515120. doi: 10.1073/pnas.2315515120. Epub 2023 Dec 20. Proc Natl Acad Sci U S A. 2023. PMID: 38117855 Free PMC article.
-
Long-term expanding human airway organoids for disease modeling.EMBO J. 2019 Feb 15;38(4):e100300. doi: 10.15252/embj.2018100300. Epub 2019 Jan 14. EMBO J. 2019. PMID: 30643021 Free PMC article.
-
Normal Calcium-Activated Anion Secretion in a Mouse Selectively Lacking TMEM16A in Intestinal Epithelium.Front Physiol. 2019 Jun 13;10:694. doi: 10.3389/fphys.2019.00694. eCollection 2019. Front Physiol. 2019. PMID: 31263421 Free PMC article.
-
Variomics screen identifies the re-entrant loop of the calcium-activated chloride channel ANO1 that facilitates channel activation.J Biol Chem. 2015 Jan 9;290(2):889-903. doi: 10.1074/jbc.M114.618140. Epub 2014 Nov 25. J Biol Chem. 2015. PMID: 25425649 Free PMC article.
References
-
- Hartzell C., Putzier I., Arreola J. (2005) Calcium-activated chloride channels. Annu. Rev. Physiol. 67, 719–758 - PubMed
-
- Eggermont J. (2004) Calcium-activated chloride channels: (un) known, (un)loved? Proc. Am. Thorac. Soc. 1, 22–27 - PubMed
-
- Yang Y. D., Cho H., Koo J. Y., Tak M. H., Cho Y., Shim W. S., Park S. P., Lee J., Lee B., Kim B. M., Raouf R., Shin Y. K., Oh U. (2008) TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 455, 1210–1215 - PubMed
-
- Caputo A., Caci E., Ferrera L., Pedemonte N., Barsanti C., Sondo E., Pfeffer U., Ravazzolo R., Zegarra-Moran O., Galietta L. J. (2008) TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science 322, 590–594 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HL73856/HL/NHLBI NIH HHS/United States
- R01 EY013574/EY/NEI NIH HHS/United States
- DK86125/DK/NIDDK NIH HHS/United States
- R01 EB000415/EB/NIBIB NIH HHS/United States
- R01 DK035124/DK/NIDDK NIH HHS/United States
- DK72517/DK/NIDDK NIH HHS/United States
- EY13574/EY/NEI NIH HHS/United States
- DK35124/DK/NIDDK NIH HHS/United States
- R01 HL073856/HL/NHLBI NIH HHS/United States
- P30 DK072517/DK/NIDDK NIH HHS/United States
- RC1 DK086125/DK/NIDDK NIH HHS/United States
- EB00415/EB/NIBIB NIH HHS/United States
- R37 DK035124/DK/NIDDK NIH HHS/United States
- R37 EB000415/EB/NIBIB NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
