

Cell motility: the integrating role of the plasma membrane
- PMID: 21833780
- PMCID: PMC3158336
- DOI: 10.1007/s00249-011-0741-0
Cell motility: the integrating role of the plasma membrane
Abstract
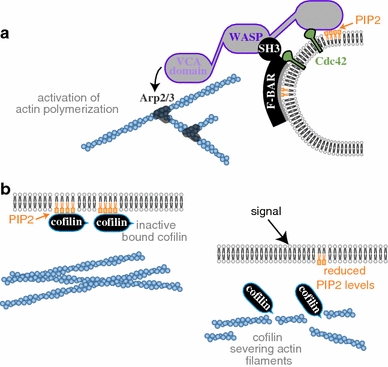
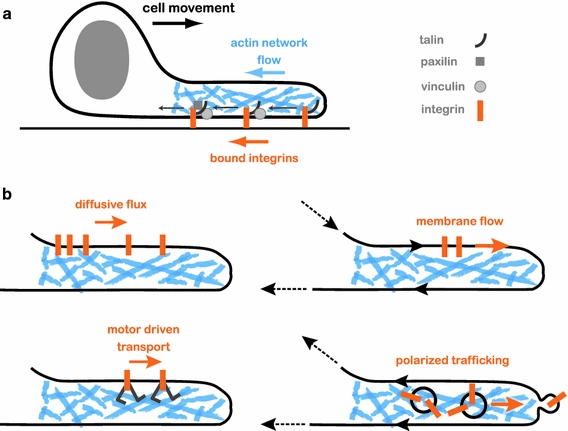
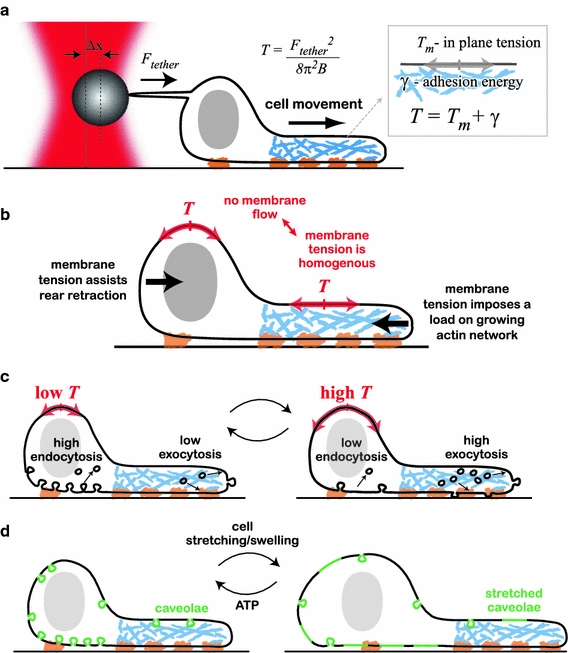
The plasma membrane is of central importance in the motility process. It defines the boundary separating the intracellular and extracellular environments, and mediates the interactions between a motile cell and its environment. Furthermore, the membrane serves as a dynamic platform for localization of various components which actively participate in all aspects of the motility process, including force generation, adhesion, signaling, and regulation. Membrane transport between internal membranes and the plasma membrane, and in particular polarized membrane transport, facilitates continuous reorganization of the plasma membrane and is thought to be involved in maintaining polarity and recycling of essential components in some motile cell types. Beyond its biochemical composition, the mechanical characteristics of the plasma membrane and, in particular, membrane tension are of central importance in cell motility; membrane tension affects the rates of all the processes which involve membrane deformation including edge extension, endocytosis, and exocytosis. Most importantly, the mechanical characteristics of the membrane and its biochemical composition are tightly intertwined; membrane tension and local curvature are largely determined by the biochemical composition of the membrane and the biochemical reactions taking place; at the same time, curvature and tension affect the localization of components and reaction rates. This review focuses on this dynamic interplay and the feedbacks between the biochemical and biophysical characteristics of the membrane and their effects on cell movement. New insight on these will be crucial for understanding the motility process.
Figures
Similar articles
-
Temporary increase in plasma membrane tension coordinates the activation of exocytosis and contraction during cell spreading.Proc Natl Acad Sci U S A. 2011 Aug 30;108(35):14467-72. doi: 10.1073/pnas.1105845108. Epub 2011 Aug 1. Proc Natl Acad Sci U S A. 2011. PMID: 21808040 Free PMC article.
-
Use the force: membrane tension as an organizer of cell shape and motility.Trends Cell Biol. 2013 Feb;23(2):47-53. doi: 10.1016/j.tcb.2012.09.006. Epub 2012 Nov 2. Trends Cell Biol. 2013. PMID: 23122885 Free PMC article. Review.
-
Cell motility through plasma membrane blebbing.J Cell Biol. 2008 Jun 16;181(6):879-84. doi: 10.1083/jcb.200802081. Epub 2008 Jun 9. J Cell Biol. 2008. PMID: 18541702 Free PMC article. Review.
-
Mechanical forces and feedbacks in cell motility.Curr Opin Cell Biol. 2013 Oct;25(5):550-7. doi: 10.1016/j.ceb.2013.06.009. Epub 2013 Jul 13. Curr Opin Cell Biol. 2013. PMID: 23860439 Review.
-
TRP Channel Trafficking.In: Liedtke WB, Heller S, editors. TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades. Boca Raton (FL): CRC Press/Taylor & Francis; 2007. Chapter 23. In: Liedtke WB, Heller S, editors. TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades. Boca Raton (FL): CRC Press/Taylor & Francis; 2007. Chapter 23. PMID: 21204515 Free Books & Documents. Review.
Cited by
-
The plasma membrane as a mechanochemical transducer.Philos Trans R Soc Lond B Biol Sci. 2019 Aug 19;374(1779):20180221. doi: 10.1098/rstb.2018.0221. Epub 2019 Jul 1. Philos Trans R Soc Lond B Biol Sci. 2019. PMID: 31431176 Free PMC article. Review.
-
Physical Plasma Membrane Perturbation Using Subcellular Optogenetics Drives Integrin-Activated Cell Migration.ACS Synth Biol. 2019 Mar 15;8(3):498-510. doi: 10.1021/acssynbio.8b00356. Epub 2019 Feb 22. ACS Synth Biol. 2019. PMID: 30764607 Free PMC article.
-
Aggregation state determines the localization and function of M1- and M23-aquaporin-4 in astrocytes.J Cell Biol. 2014 Feb 17;204(4):559-73. doi: 10.1083/jcb.201308118. Epub 2014 Feb 10. J Cell Biol. 2014. PMID: 24515349 Free PMC article.
-
Front-to-rear membrane tension gradient in rapidly moving cells.Biophys J. 2015 Apr 7;108(7):1599-1603. doi: 10.1016/j.bpj.2015.02.007. Biophys J. 2015. PMID: 25863051 Free PMC article.
-
Interactions of Functionalized Multi-Wall Carbon Nanotubes with Giant Phospholipid Vesicles as Model Cellular Membrane System.Sci Rep. 2018 Dec 20;8(1):17998. doi: 10.1038/s41598-018-36531-9. Sci Rep. 2018. PMID: 30573758 Free PMC article.
References
-
- Apodaca G. Modulation of membrane traffic by mechanical stimuli. Am J Physiol Renal Physiol. 2002;282:F179. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
