

Glucose regulates clathrin adaptors at the trans-Golgi network and endosomes
- PMID: 21832155
- PMCID: PMC3183021
- DOI: 10.1091/mbc.E11-04-0309
Glucose regulates clathrin adaptors at the trans-Golgi network and endosomes
Abstract
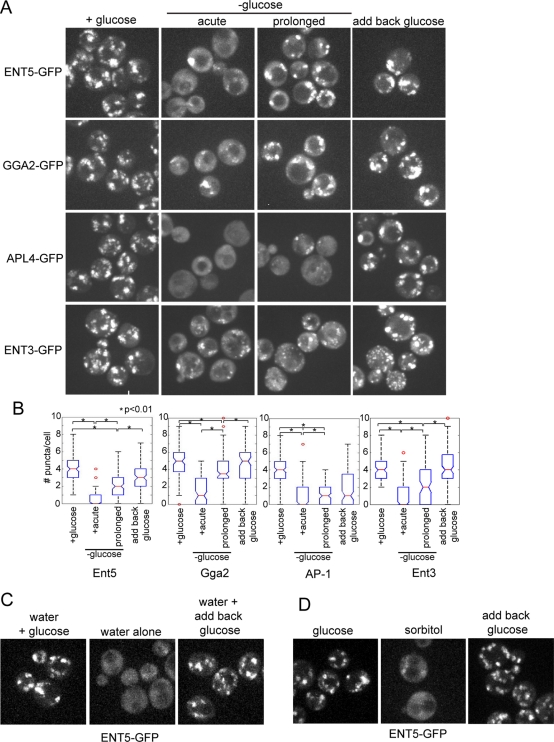
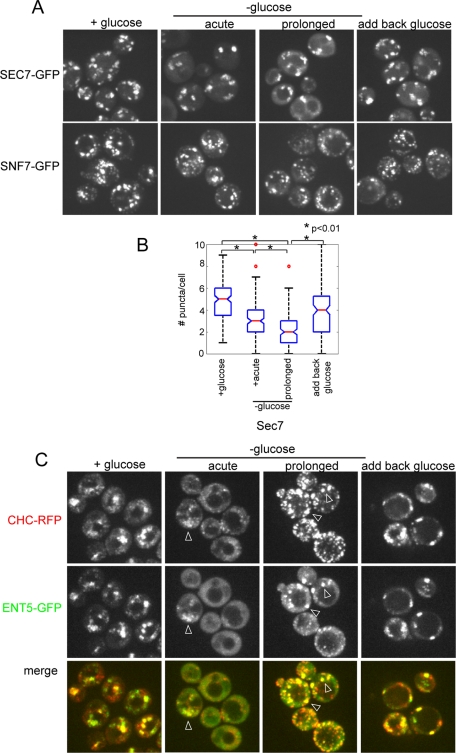
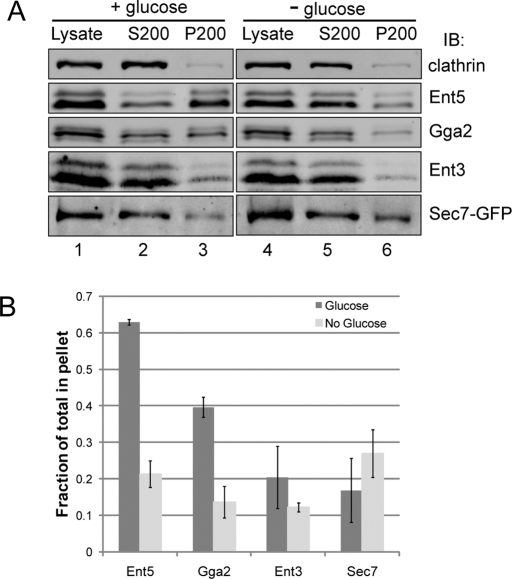
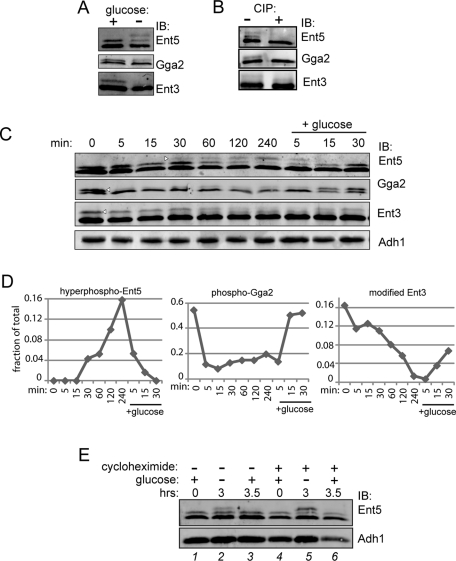
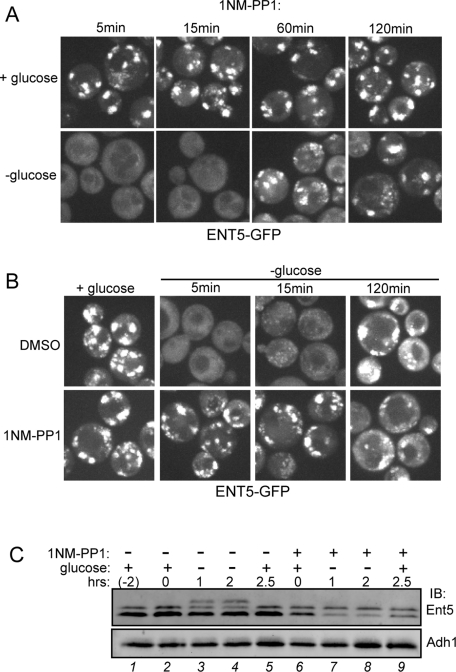
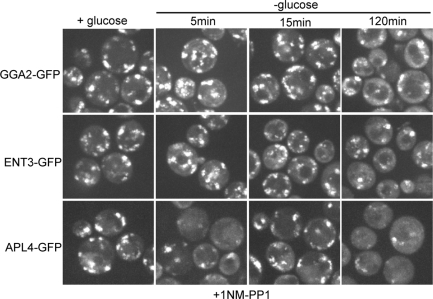
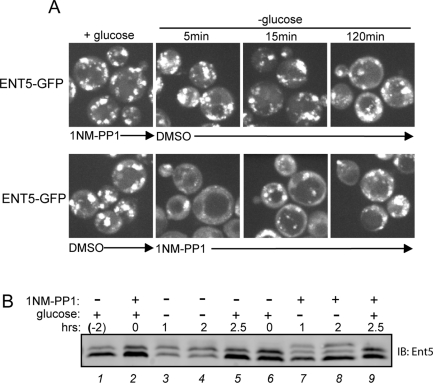
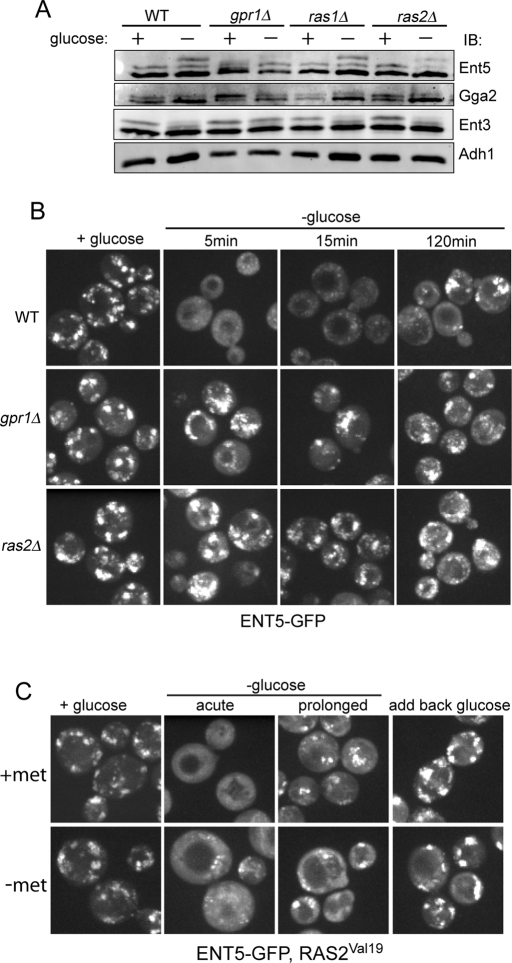
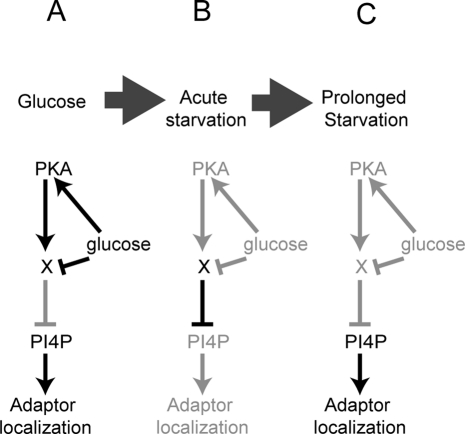
Glucose is a rich source of energy and the raw material for biomass increase. Many eukaryotic cells remodel their physiology in the presence and absence of glucose. The yeast Saccharomyces cerevisiae undergoes changes in transcription, translation, metabolism, and cell polarity in response to glucose availability. Upon glucose starvation, translation initiation and cell polarity are immediately inhibited, and then gradually recover. In this paper, we provide evidence that, as in cell polarity and translation, traffic at the trans-Golgi network (TGN) and endosomes is regulated by glucose via an unknown mechanism that depends on protein kinase A (PKA). Upon glucose withdrawal, clathrin adaptors exhibit a biphasic change in localization: they initially delocalize from the membrane within minutes and later partially recover onto membranes. Additionally, the removal of glucose induces changes in posttranslational modifications of adaptors. Ras and Gpr1 signaling pathways, which converge on PKA, are required for changes in adaptor localization and changes in posttranslational modifications. Acute inhibition of PKA demonstrates that inhibition of PKA prior to glucose withdrawal prevents several adaptor responses to starvation. This study demonstrates that PKA activity prior to glucose starvation primes membrane traffic at the TGN and endosomes in response to glucose starvation.
Figures
Similar articles
-
Energy metabolism regulates clathrin adaptors at the trans-Golgi network and endosomes.Mol Biol Cell. 2013 Mar;24(6):832-47. doi: 10.1091/mbc.E12-10-0750. Epub 2013 Jan 23. Mol Biol Cell. 2013. PMID: 23345590 Free PMC article.
-
Plasma membrane to vacuole traffic induced by glucose starvation requires Gga2-dependent sorting at the trans-Golgi network.Biol Cell. 2020 Nov;112(11):349-367. doi: 10.1111/boc.202000058. Epub 2020 Sep 3. Biol Cell. 2020. PMID: 32761633 Free PMC article.
-
Phosphoinositide-mediated clathrin adaptor progression at the trans-Golgi network.Nat Cell Biol. 2012 Feb 19;14(3):239-48. doi: 10.1038/ncb2427. Nat Cell Biol. 2012. PMID: 22344030 Free PMC article.
-
New directions for the clathrin adaptor AP-1 in cell biology and human disease.Curr Opin Cell Biol. 2022 Jun;76:102079. doi: 10.1016/j.ceb.2022.102079. Epub 2022 Apr 13. Curr Opin Cell Biol. 2022. PMID: 35429729 Free PMC article. Review.
-
Sphingolipid metabolism in trans-golgi/endosomal membranes and the regulation of intracellular homeostatic processes in eukaryotic cells.Adv Enzyme Regul. 2010;50(1):339-48. doi: 10.1016/j.advenzreg.2009.12.004. Epub 2009 Dec 23. Adv Enzyme Regul. 2010. PMID: 20005891 Review. No abstract available.
Cited by
-
Investigation of Ldb19/Art1 localization and function at the late Golgi.PLoS One. 2018 Nov 7;13(11):e0206944. doi: 10.1371/journal.pone.0206944. eCollection 2018. PLoS One. 2018. PMID: 30403748 Free PMC article.
-
mRNA localization to P-bodies in yeast is bi-phasic with many mRNAs captured in a late Bfr1p-dependent wave.J Cell Sci. 2014 Mar 15;127(Pt 6):1254-62. doi: 10.1242/jcs.139055. Epub 2014 Jan 14. J Cell Sci. 2014. PMID: 24424022 Free PMC article.
-
Glucose starvation inhibits autophagy via vacuolar hydrolysis and induces plasma membrane internalization by down-regulating recycling.J Biol Chem. 2014 Jun 13;289(24):16736-47. doi: 10.1074/jbc.M113.525782. Epub 2014 Apr 21. J Biol Chem. 2014. PMID: 24753258 Free PMC article.
-
Heterologous Expression and Auto-Activation of Human Pro-Inflammatory Caspase-1 in Saccharomyces cerevisiae and Comparison to Caspase-8.Front Immunol. 2021 Jul 14;12:668602. doi: 10.3389/fimmu.2021.668602. eCollection 2021. Front Immunol. 2021. PMID: 34335569 Free PMC article.
-
Energy metabolism regulates clathrin adaptors at the trans-Golgi network and endosomes.Mol Biol Cell. 2013 Mar;24(6):832-47. doi: 10.1091/mbc.E12-10-0750. Epub 2013 Jan 23. Mol Biol Cell. 2013. PMID: 23345590 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
