

Translational control in oocyte development
- PMID: 21690213
- PMCID: PMC3181033
- DOI: 10.1101/cshperspect.a002758
Translational control in oocyte development
Abstract
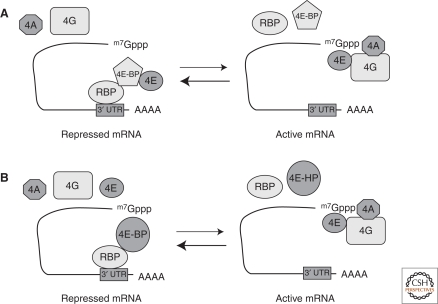
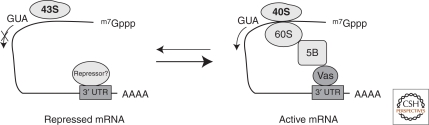
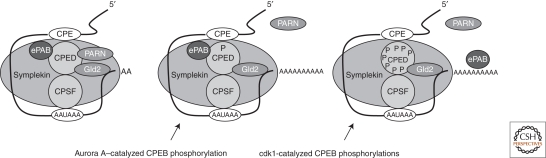
Translational control of specific mRNAs is a widespread mechanism of gene regulation, and it is especially important in pattern formation in the oocytes of organisms in which the embryonic axes are established maternally. Drosophila and Xenopus have been especially valuable in elucidating the relevant molecular mechanisms. Here, we comprehensively review what is known about translational control in these two systems, focusing on examples that illustrate key concepts that have emerged. We focus on protein-mediated translational control, rather than regulation mediated by small RNAs, as the former appears to be predominant in controlling these developmental events. Mechanisms that modulate the ability of the specific mRNAs to be recruited to the ribosome, that regulate polyadenylation of specific mRNAs, or that control the association of particular mRNAs into translationally inert ribonucleoprotein complexes will all be discussed.
Figures
Similar articles
-
Analyses of zebrafish and Xenopus oocyte maturation reveal conserved and diverged features of translational regulation of maternal cyclin B1 mRNA.BMC Dev Biol. 2009 Jan 28;9:7. doi: 10.1186/1471-213X-9-7. BMC Dev Biol. 2009. PMID: 19175933 Free PMC article.
-
Translational control by cytoplasmic polyadenylation in Xenopus oocytes.Biochim Biophys Acta. 2008 Apr;1779(4):217-29. doi: 10.1016/j.bbagrm.2008.02.002. Epub 2008 Feb 14. Biochim Biophys Acta. 2008. PMID: 18316045 Free PMC article. Review.
-
Cytoplasmic polyadenylation element (CPE)- and CPE-binding protein (CPEB)-independent mechanisms regulate early class maternal mRNA translational activation in Xenopus oocytes.J Biol Chem. 2004 Apr 23;279(17):17650-9. doi: 10.1074/jbc.M313837200. Epub 2004 Jan 29. J Biol Chem. 2004. PMID: 14752101 Free PMC article.
-
Cytoplasmic mRNA polyadenylation and translation assays.Methods Mol Biol. 2006;322:183-98. doi: 10.1007/978-1-59745-000-3_13. Methods Mol Biol. 2006. PMID: 16739724 Review.
-
Translational activation of maternally derived mRNAs in oocytes and early embryos and the role of embryonic poly(A) binding protein (EPAB).Biol Reprod. 2019 May 1;100(5):1147-1157. doi: 10.1093/biolre/ioz034. Biol Reprod. 2019. PMID: 30806655 Free PMC article. Review.
Cited by
-
Sequential Regulation of Maternal mRNAs through a Conserved cis-Acting Element in Their 3' UTRs.Cell Rep. 2018 Dec 26;25(13):3828-3843.e9. doi: 10.1016/j.celrep.2018.12.007. Cell Rep. 2018. PMID: 30590052 Free PMC article.
-
CPEB1 restrains proliferation of Glioblastoma cells through the regulation of p27(Kip1) mRNA translation.Sci Rep. 2016 May 4;6:25219. doi: 10.1038/srep25219. Sci Rep. 2016. PMID: 27142352 Free PMC article.
-
Stable intronic sequence RNA (sisRNA), a new class of noncoding RNA from the oocyte nucleus of Xenopus tropicalis.Genes Dev. 2012 Nov 15;26(22):2550-9. doi: 10.1101/gad.202184.112. Genes Dev. 2012. PMID: 23154985 Free PMC article.
-
Regulation of adult stem cell quiescence and its functions in the maintenance of tissue integrity.Nat Rev Mol Cell Biol. 2023 May;24(5):334-354. doi: 10.1038/s41580-022-00568-6. Epub 2023 Mar 15. Nat Rev Mol Cell Biol. 2023. PMID: 36922629 Free PMC article. Review.
-
A Role of PI3K/Akt Signaling in Oocyte Maturation and Early Embryo Development.Cells. 2023 Jul 12;12(14):1830. doi: 10.3390/cells12141830. Cells. 2023. PMID: 37508495 Free PMC article. Review.
References
-
- Barnard DC, Ryan K, Manley JL, Richter JD 2004. Symplekin and xGLD-2 are required for CPEB-mediated cytoplasmic polyadenylation. Cell 119: 641–651 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases