

A critical function for Ser-282 in cardiac Myosin binding protein-C phosphorylation and cardiac function
- PMID: 21597010
- PMCID: PMC3132348
- DOI: 10.1161/CIRCRESAHA.111.242560
A critical function for Ser-282 in cardiac Myosin binding protein-C phosphorylation and cardiac function
Abstract
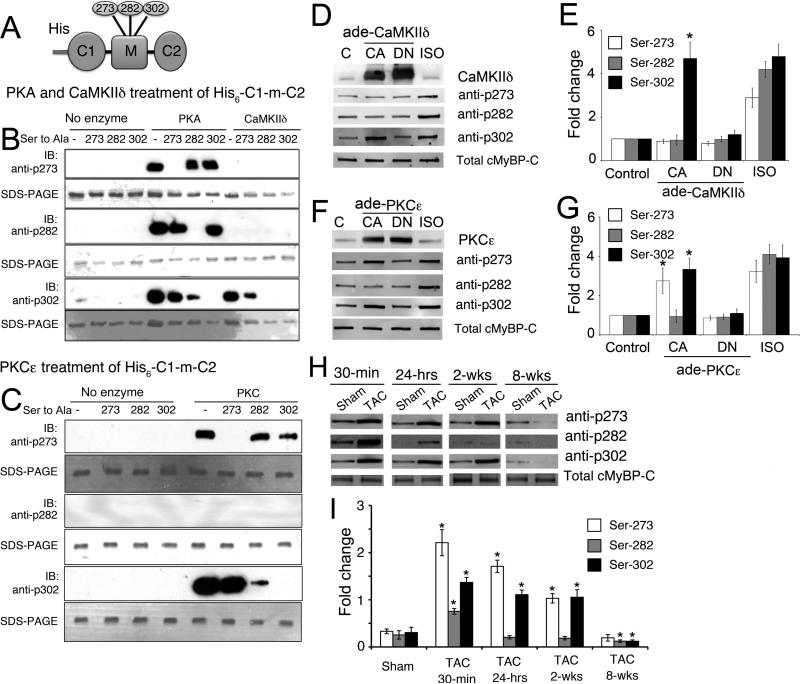
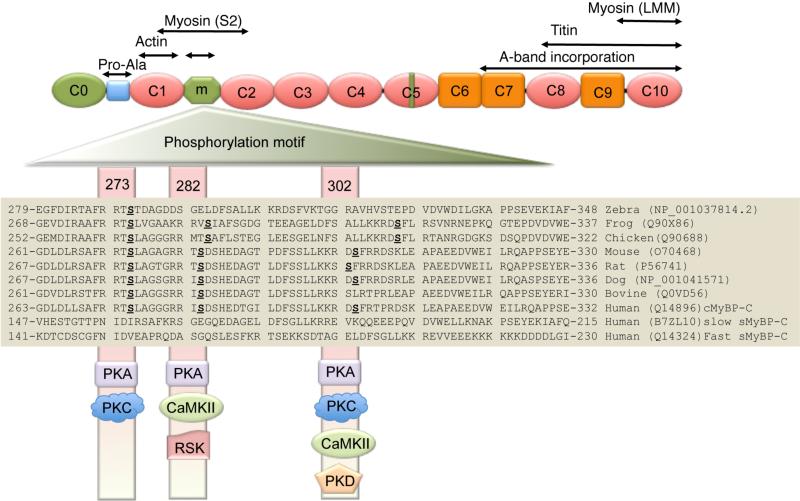
Rationale: Cardiac myosin-binding protein-C (cMyBP-C) phosphorylation at Ser-273, Ser-282, and Ser-302 regulates myocardial contractility. In vitro and in vivo experiments suggest the nonequivalence of these sites and the potential importance of Ser-282 phosphorylation in modulating the protein's overall phosphorylation and myocardial function.
Objective: To determine whether complete cMyBP-C phosphorylation is dependent on Ser-282 phosphorylation and to define its role in myocardial function. We hypothesized that Ser-282 regulates Ser-302 phosphorylation and cardiac function during β-adrenergic stimulation.
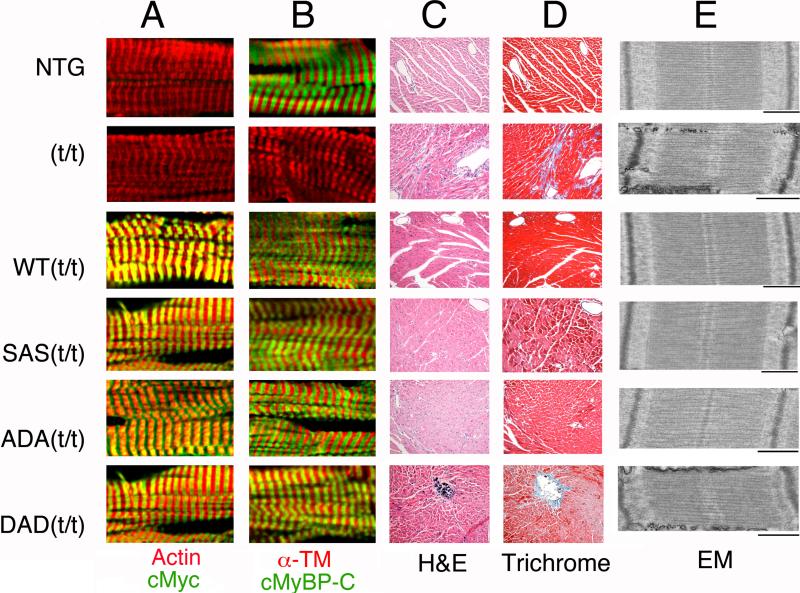
Methods and results: Using recombinant human C1-M-C2 peptides in vitro, we determined that protein kinase A can phosphorylate Ser-273, Ser-282, and Ser-302. Protein kinase C can also phosphorylate Ser-273 and Ser-302. In contrast, Ca(2+)-calmodulin-activated kinase II targets Ser-302 but can also target Ser-282 at nonphysiological calcium concentrations. Strikingly, Ser-302 phosphorylation by Ca(2+)-calmodulin-activated kinase II was abolished by ablating the ability of Ser-282 to be phosphorylated via alanine substitution. To determine the functional roles of the sites in vivo, three transgenic lines, which expressed cMyBP-C containing either Ser-273-Ala-282-Ser-302 (cMyBP-C(SAS)), Ala-273-Asp-282-Ala-302 (cMyBP-C(ADA)), or Asp-273-Ala-282-Asp-302 (cMyBP-C(DAD)), were generated. Mutant protein was completely substituted for endogenous cMyBP-C by breeding each mouse line into a cMyBP-C null (t/t) background. Serine-to-alanine substitutions were used to ablate the abilities of the residues to be phosphorylated, whereas serine-to-aspartate substitutions were used to mimic the charged state conferred by phosphorylation. Compared to control nontransgenic mice, as well as transgenic mice expressing wild-type cMyBP-C, the transgenic cMyBP-C(SAS(t/t)), cMyBP-C(ADA(t/t)), and cMyBP-C(DAD(t/t)) mice showed no increases in morbidity and mortality and partially rescued the cMyBP-C((t/t)) phenotype. The loss of cMyBP-C phosphorylation at Ser-282 led to an altered β-adrenergic response. In vivo hemodynamic studies revealed that contractility was unaffected but that cMyBP-C(SAS(t/t)) hearts showed decreased diastolic function at baseline. However, the normal increases in cardiac function (increased contractility/relaxation) as a result of infusion of β-agonist was significantly decreased in all of the mutants, suggesting that competency for phosphorylation at multiple sites in cMyBP-C is a prerequisite for normal β-adrenergic responsiveness.
Conclusions: Ser-282 has a unique regulatory role in that its phosphorylation is critical for the subsequent phosphorylation of Ser-302. However, each residue plays a role in regulating the contractile response to β-agonist stimulation.
Figures


Similar articles
-
Phosphoregulation of Cardiac Inotropy via Myosin Binding Protein-C During Increased Pacing Frequency or β1-Adrenergic Stimulation.Circ Heart Fail. 2015 May;8(3):595-604. doi: 10.1161/CIRCHEARTFAILURE.114.001585. Epub 2015 Mar 4. Circ Heart Fail. 2015. PMID: 25740838 Free PMC article.
-
Functional dissection of myosin binding protein C phosphorylation.J Mol Cell Cardiol. 2013 Nov;64:39-50. doi: 10.1016/j.yjmcc.2013.08.006. Epub 2013 Aug 31. J Mol Cell Cardiol. 2013. PMID: 24001940 Free PMC article.
-
Acceleration of crossbridge kinetics by protein kinase A phosphorylation of cardiac myosin binding protein C modulates cardiac function.Circ Res. 2008 Oct 24;103(9):974-82. doi: 10.1161/CIRCRESAHA.108.177683. Epub 2008 Sep 18. Circ Res. 2008. PMID: 18802026 Free PMC article.
-
Phosphorylation and function of cardiac myosin binding protein-C in health and disease.J Mol Cell Cardiol. 2010 May;48(5):866-75. doi: 10.1016/j.yjmcc.2009.11.014. Epub 2009 Dec 3. J Mol Cell Cardiol. 2010. PMID: 19962384 Free PMC article. Review.
-
Species-specific differences in the Pro-Ala rich region of cardiac myosin binding protein-C.J Muscle Res Cell Motil. 2009 Dec;30(7-8):303-6. doi: 10.1007/s10974-010-9207-8. Epub 2010 Mar 9. J Muscle Res Cell Motil. 2009. PMID: 20217194 Free PMC article. Review.
Cited by
-
Cardiac myosin binding protein C phosphorylation in cardiac disease.J Muscle Res Cell Motil. 2012 May;33(1):43-52. doi: 10.1007/s10974-011-9280-7. Epub 2011 Nov 30. J Muscle Res Cell Motil. 2012. PMID: 22127559 Free PMC article. Review.
-
Oxidative stress and sarcomeric proteins.Circ Res. 2013 Jan 18;112(2):393-405. doi: 10.1161/CIRCRESAHA.111.300496. Circ Res. 2013. PMID: 23329794 Free PMC article. Review.
-
Hypertrophic cardiomyopathy in MYBPC3 carriers in aging.J Cardiovasc Aging. 2024 Jan;4(1):9. doi: 10.20517/jca.2023.29. Epub 2024 Jan 11. J Cardiovasc Aging. 2024. PMID: 38406555 Free PMC article.
-
Modulation of myosin by cardiac myosin binding protein-C peptides improves cardiac contractility in ex-vivo experimental heart failure models.Sci Rep. 2022 Mar 14;12(1):4337. doi: 10.1038/s41598-022-08169-1. Sci Rep. 2022. PMID: 35288601 Free PMC article.
-
Biomarkers for Heart Failure Prognosis: Proteins, Genetic Scores and Non-coding RNAs.Front Cardiovasc Med. 2020 Nov 23;7:601364. doi: 10.3389/fcvm.2020.601364. eCollection 2020. Front Cardiovasc Med. 2020. PMID: 33330662 Free PMC article. Review.
References
-
- Thom T, Haase N, Rosamond W, Howard VJ, Rumsfeld J, Manolio T, Zheng ZJ, Flegal K, O'Donnell C, Kittner S, Lloyd-Jones D, Goff DC, Jr., Hong Y, Adams R, Friday G, Furie K, Gorelick P, Kissela B, Marler J, Meigs J, Roger V, Sidney S, Sorlie P, Steinberger J, Wasserthiel-Smoller S, Wilson M, Wolf P. Heart disease and stroke statistics--2006 update: a report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation. 2006;113:e85–151. - PubMed
-
- Liao R, Wang CK, Cheung HC. Coupling of calcium to the interaction of troponin I with troponin C from cardiac muscle. Biochemistry. 1994;33:12729–12734. - PubMed
-
- Houser SR, Piacentino V, 3rd, Weisser J. Abnormalities of calcium cycling in the hypertrophied and failing heart. J Mol Cell Cardiol. 2000;32:1595–1607. - PubMed
-
- El-Armouche A, Boknik P, Eschenhagen T, Carrier L, Knaut M, Ravens U, Dobrev D. Molecular determinants of altered Ca2+ handling in human chronic atrial fibrillation. Circulation. 2006;114:670–680. - PubMed
-
- Decker RS, Decker ML, Kulikovskaya I, Nakamura S, Lee DC, Harris K, Klocke FJ, Winegrad S. Myosin binding protein C phosphorylation, myofibril structure, and contractile function during low-flow ischemia. Circulation. 2005;111:906–912. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 HL059408-11/HL/NHLBI NIH HHS/United States
- P01 HL059408-12/HL/NHLBI NIH HHS/United States
- P01 HL069779/HL/NHLBI NIH HHS/United States
- P01 HL059408/HL/NHLBI NIH HHS/United States
- R01HL087862/HL/NHLBI NIH HHS/United States
- P01 HL059408-13/HL/NHLBI NIH HHS/United States
- R01 HL087862-03/HL/NHLBI NIH HHS/United States
- P50HL074728/HL/NHLBI NIH HHS/United States
- P01HL69799/HL/NHLBI NIH HHS/United States
- R01 HL105826-01/HL/NHLBI NIH HHS/United States
- R37 HL030077/HL/NHLBI NIH HHS/United States
- P50HL077101/HL/NHLBI NIH HHS/United States
- R01 HL105826/HL/NHLBI NIH HHS/United States
- P50 HL074728/HL/NHLBI NIH HHS/United States
- R01 HL087862/HL/NHLBI NIH HHS/United States
- R01 HL105826-02/HL/NHLBI NIH HHS/United States
- P01 HL069779-10/HL/NHLBI NIH HHS/United States
- P01HL059408/HL/NHLBI NIH HHS/United States
- R01HL105826/HL/NHLBI NIH HHS/United States
- P50 HL077101/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
